VetBact
Veterinärmedicinsk bakteriologi: information om betydelsefulla arter
Veterinärmedicinsk bakteriologi
.")
Strukturformel för ATP. De tre fosfatgrupperna benämns alfa, beta och gamma räknat från ribos-delen. Kvävebasen (adenin) bildar tillsammans med sockerdelen (ribos) molekylen adenosin. Har adenosin bara en fosfatgrupp, så kallas den adenosinmonofosfat (AMP). - Klicka på bilden för att förstiora den.
Adenosintrifosfat (ATP) är en nukleotid (nukleosidtrifosfat), som består av en kvävebas (adenin), en sockermolekyl (ribos) och en trifosfatgrupp. ATP utgör den viktigaste energiformen för alla organismer och kan sägas vara cellens energivaluta. Anledningen till detta är att ATP innehåller energirika bindningar mellan fosfatgrupperna och den kemiska energin i dessa bindningar kan utvinnas när en sådan grupp spjälkas av från ATP och då bildas det ADP (adenosindifosfat) eller från ADP till AMP (adenosinmonofosfat). Cellen kan inte lagra ATP, utan syntesen av ATP måste hela tiden regleras i förhållande till behovet. ATP är också en byggsten vid syntes av nukleinsyra.
Energi krävs för syntes av ATP från ADP och fosfat. Den energin kan bildas vid glykolys, under Krebscykeln och genom elektrontransportkedjan. Vissa intracellulära bakterier (t.ex. medlemmar av familjen Chlamydiaceae) kan inte syntetisera den mängd av ATP, som krävs för cellens olika funktioner, utan måste därför parasitera på värdcellens ATP-syntes.
Man kan dela in de energikrävande processerna, som utnyttjar ATP, i en bakteriecell i tre huvudgrupper:
Den tysk-brittiska biokemisten Hans Krebs fick nobelpris i fyiologi eller medicin 1953 för att han utredde och presenterade citronsyracykelns viktigaste delar på 1930-talet.
Uppdaterad: 2019-10-17.
Med adhesion i bakteriologiska sammanhang menar man bakteriers förmåga till vidhäftning på ytor (t.ex. vävnader eller främmande material d.v.s. proteser), som kan vara mer eller mindre specifik. Kolonisation av bakterier till en yta och bildandet av en biofilm börjar med adhesion till ytan. Om ytan utgörs av ett inert främmande material är adhesionen oftast ospecifik.
För att specifik adhesion ska kunna äga rum, måste det finnas en receptor på ytan och en ligand på bakterien, som kan binda till ytan. Bakterier använder s.k. adhesiner som ligander för att kunna fästa till olika receptorer på celler i värddjurets vävnad. Adhesiner utgörs av proteiner eller polysackarider och även dess receptorer kan utgöras av proteiner eller polysackarider. Adhesiner är viktiga virulensfaktorer eftersom de bidrar till att bakterier kan kolonisera olika vävnader. Många patogena bakterier kan producera ett stort utbud av olika adhesiner, som uttrycks under olika faser av infektionen. Inhibering av ett av dessa adhesiner resulterar oftast i att bakterien förlorar sin patogenicitet. Försök att utnyttja anti-adhesinantikroppar, som vaccin har i vissa fall varit framgångsrika i modelsystem.
![Adhesion (Vidhäftning) [under utarbetande]](/images/terms/large/230517_Sv_v2.jpg "Klicka på bilden för att förstora den (öppnas i ett nytt fönster).")
Ett välkaraktäriserat bakteriellt adhesin är FimH, som är bundet till pili och produceras av uropatogena Escherichia coli (UPEC). FimH binder till mannos och CD48 är en typ av humant immunoglobulin, som är glykosylerat och innehåller mannosrester. TLR4 är en medlem i den s.k. toll-like-receptor- familjen, som utgörs av humana membranproteiner och är viktiga för den medfödda immuniteten. Bilden är anpassad från Wikipedia, Bacterial adhesion. - Klicka på bilden för att förstora den.
Fimbrier (= vanliga pili) fungerar ofta som adhesiner och ibland kan en speciell polypeptid, som sitter i spetsen på fimbrien, utgöra själva liganden. Membranproteiner kan också fungera som adhesiner.
Många bakterier producerar extracellulära polysackarider, som utgör ospecifika adhesionsfaktorer. Den extracellulära polysackariden poly-N-acetylglukosamin (PNAG) fungerar t.ex. som adhesionsfaktor för vissa stafylokocker och gör att de kan fastna på vissa abiotiska (icke-levande) ytor och där bilda en biofilm.
Di Martino, P. 2018. Bacterial adherence: Much more than a bond. AIMS Microbiol. 4(3): 563-566.
Uppdaterad: 2023-05-31.
Anaeroba bakterier är bakterier, som inte kan utnyttja syre i sin metabolism utan förgiftas och dör av syrgas. Man brukar också säga att sådana bakterier är strikt anaeroba (= obligat anaeroba) för att skilja dem från syretoleranta bakterier och från fakultativt anaeroba (= fakultativt aeroba) bakterier. Syretoleranta bakterier använder ej syre i sin metabolism, men förgiftas ej av syrgas utan kan leva och föröka sig i närvaro av syrgas, åtminstone under en viss tid. Fakultativt anaeroba bakterier förgiftas inte heller av syrgas och kan även ställa om sin metabolism, så att de i närvaro av syrgas utnyttjar syre i metabolismen, men i frånvaro av syrgas kan de utvinna energi på andra sätt (t.ex. genom fermentation eller anaerob respiration).
 Syremolekylen består av två atomer syre, vilket gör molekylen stabil eftersom de två atomerna då kan dela på ett elektronpar i det yttre elektronskalet (se Fig. A). I närvaro av syrgas (O2) bildas i en vattenlösning (t.ex. i en cell) alltid små mängder av väteperoxid (H2O2) och superoxidradikaler genom en jämviktsreaktion. Superoxidradikalen (se Fig. B) brukar skrivas O2·-, för att visa att den består av en syremolekyl, som har tagit upp en extra elektron (·), vilket gör den negativt laddad. Väteperoxid och framför allt superoxidradikalen är mycket giftiga för celler eftersom de är synnerligen reaktiva och kan påverka en mängd olika substanser, t.ex. oxidera omättade fettsyror, vilket leder till s.k. oxidativ stress. För att kunna överleva i syrehaltig miljö, måste celler därför ha enzymer, som kan metabolisera (avgifta) väteperoxid och superoxidradikalen.
Syremolekylen består av två atomer syre, vilket gör molekylen stabil eftersom de två atomerna då kan dela på ett elektronpar i det yttre elektronskalet (se Fig. A). I närvaro av syrgas (O2) bildas i en vattenlösning (t.ex. i en cell) alltid små mängder av väteperoxid (H2O2) och superoxidradikaler genom en jämviktsreaktion. Superoxidradikalen (se Fig. B) brukar skrivas O2·-, för att visa att den består av en syremolekyl, som har tagit upp en extra elektron (·), vilket gör den negativt laddad. Väteperoxid och framför allt superoxidradikalen är mycket giftiga för celler eftersom de är synnerligen reaktiva och kan påverka en mängd olika substanser, t.ex. oxidera omättade fettsyror, vilket leder till s.k. oxidativ stress. För att kunna överleva i syrehaltig miljö, måste celler därför ha enzymer, som kan metabolisera (avgifta) väteperoxid och superoxidradikalen.
 Strikt anaeroba bakterier saknar alla enzymer, som kan metabolisera väteperoxid och superoxidradikaler. Övriga bakterier har superoxiddismutas (SODM), som omvandlar superoxidradikaler till väteperoxid och syrgas (se Fig. C). Aeroba och de flesta fakultativt anaeroba bakterierna har katalas, som omvandlar väteperoxid till vatten och syrgas (se Fig. C). Många syretoleranta anaeroba bakterier har ett peroxidas, som omvandlar väteperoxid till vatten med hjälp av NADH2 (se Fig. C; klicka på bilden för att förstora den).
Strikt anaeroba bakterier saknar alla enzymer, som kan metabolisera väteperoxid och superoxidradikaler. Övriga bakterier har superoxiddismutas (SODM), som omvandlar superoxidradikaler till väteperoxid och syrgas (se Fig. C). Aeroba och de flesta fakultativt anaeroba bakterierna har katalas, som omvandlar väteperoxid till vatten och syrgas (se Fig. C). Många syretoleranta anaeroba bakterier har ett peroxidas, som omvandlar väteperoxid till vatten med hjälp av NADH2 (se Fig. C; klicka på bilden för att förstora den).
Uppdaterad: 2017-11-09.
.")
Anaerob odling med hjälp av kommersiella gasgenererande kuvert i lufttät anaerobklocka. I klocka finns en indikator strip som blir vit vid anaerob miljö. - Klicka på bilden för att förstora den.
Vissa patogena bakterier är anaeroba och måste odlas i syrefri atmosfär. Vilken metoden som används beror på hur anaerob bakterien som ska odlas är. Det finns strikt anaeroba bakterier, som inte alls tål syre, och det finns syretoleranta anaeroba bakterier, som tål syre under kortare perioder.
Om man jobbar med strikta anaerober eller om man har stora mängder av prover, som man ska odla ifrån, kanske man föredrar ett s.k. anaerobskåp där man arbetar med gummihandskar förseglade till skåpet. Anaerobskåpet håller konstant temperatur och atmosfären kan t.ex bestå av kvävgas (N2) och koldioxid (CO2).
Om man jobbar med syretoleranta anaerober kanske man fördrar en s.k. anaerobklocka där man med hjälp av kemikalier absorberar syre (O2) och genererar CO2. Det finns kommersiella system för detta ändamål. Anaerobklockan kan ställas in i ett värmeskåp vid inkuberingen.
Uppdaterad: 2020-02-26.
Ett antibiotikum (pl. antibiotika) är en substans, som selektivt dödar (baktericida) eller hämmar (bakteriostatiska) tillväxten av bakterier. Nedan följer en sammanställning av olika antibiotika, som används inom veterinärmedicinen. Det är dock inte säkert att alla antibiotika, som nämns nedan, finns registrerade för användning på djur i Sverige. När det gäller behandlingar vid specifika sjukdomar på olika djurslag hänvisar vi till Sveriges Veterinärförbunds (SVFs) Antibiotikapolicy. Där kan man hitta generella riktlinjer för antibiotikabehandling av djur samt specifika riktlinjer när det gäller smådjur, häst och lantbrukets djur.
Bakterier kan vara resistenta mot vissa antibiotika och man brukar tala om naturlig eller förvärvad resistens. Naturlig resistens innebär att en bakterie är resistent mot ett antibiotikum tack vare sin naturliga egenskap. Mykoplasmer är t.ex. naturligt resistenta mot antibiotika, som hämmar cellväggssyntesen, eftersom de saknar cellväg. Förvärvad resistens innebär att en bakterie, som från början var känslig för ett visst antibiotikum, har utvecklat resistens (genom selektion) på grund av att den har exponerats för detta antibiotikum. Antibiotikaresistens är ett mycket stort och, tyvärr, växande problem inom all sjukvård på grund av en tidigare alltför frikostig förskrivning av antibiotika, vilket olyckligtvis gäller fortfarande i vissa länder och även inom EU. Från Statens Veterinärmedicinska Anstalts (SVAs) webbplats kan man ladda ner de s.k. SVARM- och SVARM/SWEDRES-rapporterna, som publiceras årligen av Folkhälsomyndigheten och SVA. SVARM- och SVARM/SWEDRES-rapporterna ger en sammanfattning av antibiotikaförbrukningen och resistensläget i Sverige inom både veterinär- och humanmedicin. Bakteriers förmåga att överleva i närvaro av ett antibiotikum kan undersökas genom en s.k. resistensbestämning. Resultatet av en resistensbestämning uttrycks i form av ett MIC-värde (Minimum Inhibitory Concentration), som har sorten mg/l (= µg/ml). För att underlätta tolkningen av ett MIC-värde brukar man klassificera ett bakterieisolat som känsligt (S), intermediärt (I) eller resistent (R) beroende på hur MIC-värdet ligger i förhållande till bakteriens "normala" MIC-fördelning (se nedan).
Antibiotika brukar indelas i olika grupper beroende på deras verkningsmekanismer och kemiska struktur. De antibiotika, som finns nämnda i SVFs policydokument, är skrivna i fet stil när de först förekommer i texten nedan.
A. Beta-laktamer, där den s.k. beta-laktamringen ingår i strukturen.
1. Penicilliner (t.ex. aminopenicillin, amoxicillin, ampicillin, bensylpenicillin, fenoxymetylpenicillin, kloxacillin och meticillin).
2. Cefalosporiner (t.ex. cefalexin, cefalotin, cefotaxim och cefepim). Cefalosporiner ska användas mycket restriktivt för behandling av djur eftersom de också används för behandling av livshotande sjukdomar hos människa. Användning av tredje och fjärde generationens cefalosporiner är reglerad i författningen SJVFS 2013:42, Saknr D9.
3. Cefamyciner (latamoxef) liknar cefalosporiner.
4. Karbapenemer (t.ex. imipenem, meropenem och aztreonam) är idag förbjudna att använda för behandling av djur eftersom de också används för behandling av livshotande sjukdomar hos människa (författning SJVFS 2013:42, Saknr D9, bilaga).
B. Andra hämmare av cellväggssyntesen.
1. Glykopeptider (t.ex. vankomycin och teikoplanin) blockerar transpeptideringen genom att binda till -D-alanin-D-alanin- i peptidoglykanens sidopeptid. Glykopeptider är idag förbjudna att använda för behandling av djur eftersom de också används för behandling av livshotande sjukdomar hos människa (författning SJVFS 2013:42, Saknr D9, bilaga).
Vid syntesen av cellväggen hos bakterier ingår bl.a. att s.k. transpeptidas, som binder till dipeptiden -ala-ala- och kopplar ihop peptiderna som ska ingå i cellväggen, till ett nätverk. Beta-laktamringen är en strukturanalog till -ala-ala- och kan därför konkurrera ut -ala-ala- vilket innebär att det inte kan bildas någon stabil cellvägg. Mutationer i transpeptidaset eller andra s.k. penicillinbindande proteiner (PBP) kan leda till att penicillinet inte längre binder och bakterien blir därför resistent.
Vissa bakterier har naturligt ett kromosom- eller plasmidkodat penicillinas, d.v.s. ett enzym som hydrolyserar beta-laktamringen. I närvaro av beta-laktamer kan dessa bakterier stimuleras till att öka produktionen av beta-laktamas och om beta-laktamasgenen är plasmidkodad, kan den överföras till andra bakterier. Klavulansyra, som är en strukturanalog till penicilliner, kan inhibera penicillinaser genom att blockera deras aktiva centra. Klavulansyra är speciellt aktiv mot plasmidburna beta-laktamaser och används terapeutiskt. Klavulansyra är inte ett antibiotikum i sig, eftersom det endast är verksamt i kombination med vissa beta-laktamer.
A. Fluorokinoloner (t.ex. ciprofloxacin, difloxacin, enrofloxacin, ibafloxacin, marbofloxacin och orbifloxacin)
hämmar enzymerna DNA-gyras och topoisomeras, som behövs för DNA-replikationen. Fluorokinoloner ska användas mycket restriktivt för behandling av djur eftersom de också används för behandling av livshotande sjukdomar hos människa. Användning av tredje och fjärde generationens fluorokinoloner är reglerad i författningen SJVFS 2013:42, Saknr D9.
B. Rifampicin hämmar DNA-beroende RNA-polymeras, vilket inhiberar proteinsyntesen genom att syntesen av mRNA hämmas. Rifampicin får endast användas för behandling av Rhodoccus-infectioner (författning SJVFS 2013:42, Saknr D9, bilaga).
C. Nitroimidazoler (t.ex. dimetridazol, metronidazol och ronidazol) reagerar med nukleoproteiner i anaerob miljö, vilket i sin tur resulterar i brott på DNA-spiralen. Metronidazol är ett av de få antibiotika, som är verksamt mot Clostridium difficile-orsakade diarréer hos människa och ska därför endast undantagsvis användas till djur.
Resistens mot fluorokinoloner kan uppkomma genom mutation(er) i generna för DNA-gyras och/eller topoisomeras, vilket göra att fluorokinolonerna inte längre kan blockera dessa enzymer.
A. Aminoglykosider (t.ex. gentamicin, neomycin, streptomycin och dihydrostreptomycin) binder till 30S subenheten på bakterieribosomer och orsakar felläsning av den genetiska koden.
B. Tetracykliner (klortetracyklin, oxitetracyklin och doxycyklin) interfererar med aminoacylerade tRNA-kedjor och förhindrar bindning av aminosyror till den växande polypeptidkedjan. Detta leder till att proteinsyntesen inhiberas.
C. Kloramfenikol (kloromycetin) hämmar enzymet peptidyltransferas, som kopplar ihop aminosyror med den växande polypeptidkedjan. Detta leder till att proteinsyntesen inhiberas.
D. Makrolider (t.ex. tylosin och erytromycin) och linkosamider (t.ex. linkomycin och klindamycin) binder till 23S rRNA i bakteriens ribosom och förhindrar translokationen längs med mRNA-kedjan. Ketolider hör till gruppen makrolider, men de har ett bredare bakteriellt spektrum. Azalider (t.ex azitromycin) och streptograminer hör också till gruppen makrolider.
E. Pleuromutiliner (t.ex. valnemulin och tiamulin) binder till peptidyltransferaset vid 50S subenheten och förhindrar därmed att aminosyror kopplas ihop med den växande polypeptidkedjan.
F. Fusidinsyra är ett steroidderivat, som interfererar med frisättning av elongeringsfaktorn EF-G när aminosyror kopplas till den växande polypeptidkedjan.
G. Oxazolidinoner binder till 23S rRNA i bakteriens ribosom och förhindrar translokationen längs med mRNA-kedjan. Oxazolidinoner är idag förbjudna att använda för behandling av djur eftersom de också används för behandling av livshotande sjukdomar hos människa (författning SJVFS 2013:42, Saknr D9, bilaga).
Aminoglykosidresistens kan uppkomma genom mutationer i ribosomproteiner, så att inte makroliden kan binda till ribosomen längre. Vissa bakterier bär på plasmider, som kodar för aminoglykosidmodifierande enzymer. Sådana enzymer inaktiverar aminoglykosider och kan lätt spridas i en bakteriepopulation.
Linkosamid- och makrolidresistens kan uppkomma genom att bakterien kan metylera en viss nukleotid i 23S rRNA eller genom mutationer i 23S rRNA-genen, vilket gör att makroliden inte längre binder till ribosomen.
A. Polypeptidantibiotika (t.ex. polymyxin B och kolistin) påverkar cellmembranets permeabilitet genom att binda till LPS och interferera med fosfolipider. Detta gör att essentiella metaboliter läcker ut ur baktericellen.
B. Cykliska lipopeptider (t.ex. daptomycin) binder till bakteriernas cellmembran, vilket då depolariseras. Detta leder till inhibering av DNA-, RNA- och proteinsyntes.
A. Folsyraantagonister
1. Sulfonamider (t.ex. sulfa och sulfadoxin) hämmar syntes av folsyra genom att konkurrera ut strukturanalogen paraaminobenzoesyra. Folsyra är ett koenzym, som krävs vid syntesen av vissa aminosyror och nukleotider och som eukaryota celler kan ta upp från sin omgivning.
2. Trimetroprim hämmar enzymet dihydrofolatreduktas. Sulfa och trimetroprim kombineras ofta för att motverka uppkomst av resistens.
B. Furantoiner (t.ex. nitrofurantoin och furadantin) omvandlas av bakteriella flavoproteiner till reaktiva intermediat, som inhiberar bakteriernas energimetabolism samt syntes av DNA, RNA, proteiner och cellvägg. Genom att furantoiner påverkar så många olika steg i bakteriernas metabolism, så uppkommer resistens inte så lätt och därför används dessa mot urinvägsinfektioner på framför allt hund för att minska användningen av aminopenicilliner där resistens lättare uppkommer. Resistens mot furantoiner förekommer men är ovanlig.
Punktmutationer i generna för en del av de enzymer som ingår i syntesen av folsyra kan leda till resistens genom att dessa modifierade enzymer inte längre blockeras. Enterokocker är naturligt resistenta mot sulfonamider eftersom de tar upp folsyra från omgivningen.
Genom minskad permeabilitet eller ökat utflöde kan bakterier utveckla resistensmekanismer, som är verksamma mot olika typer av antibiotika. Minskat upptag kan bero på mutationer i poriner och ökat utflöde kan bero på mutationer i s.k. effluxpumpar. Effluxpumpar är proteinkomplex, som kan pumpa ut substanser ur cellen genom aktiv transport. Oftast har effluxpumparna naturliga substrat, som gallsalter, adhesiner, toxiner och andra proteiner.
ESBL, är en förkortning av "Extended Spectrum Beta-Lactamases", vilket betyder beta-laktamas med utvidgat spektrum (kan hydrolysera olika typer av beta-laktamer). ESBL används ibland lite slarvigt för en grupp av multiresistenta bakterier, men är egentligen en beteckning på en grupp av enzymer, som finns hos vissa multiresistenta bakterier. ESBL ger resistens mot penicilliner och många cefalosporiner. ESBL är oftast plasmidkodat och kan överföras mellan gramnegativa bakterier.
ESBL-CARBA (eller ESBLCARBA), är beteckningen för en resistensmekanism hos tarmbakterier, som orsakar resistens mot de flesta penicilliner, cefalosporiner och karbapenemer. I Sverige är fynd av ESBLCARBA på djur anmälningspliktigt till Länsstyrelse och Jordbruksverket.
MRS, står för MeticillinResistenta Stafylokocker. MRS är en beteckning för stafylokocker i allmänhet, som är resistenta mot alla penicilliner (t.ex. meticillin) och cefalosporiner. I Sverige är fynd av MRS på djur anmälningspliktigt till Länsstyrelse och Jordbruksverket.
MRSA, står för MeticillinResistenta Staphylococcus Aureus. MRSA är en beteckning på Staphylococcus aureus-stammar, som är resistenta mot alla penicilliner (t.ex. meticillin) och cefalosporiner. I Sverige är fynd av MRSA på djur anmälningspliktigt till Länsstyrelse och Jordbruksverket.
MRSP, står för MeticillinResistenta Staphylococcus Pseudintermedius. MRSP är en beteckning på Staphylococcus pseudintermedius-stammar, som är resistenta mot alla penicilliner (t.ex. meticillin) och cefalosporiner. I Sverige är fynd av MRSP på djur anmälningspliktigt till Länsstyrelse och Jordbruksverket.
VRE, står för VancomycinResistenta Enterokocker.
Mer information om antibiotikaresistensläget och antibiotikakonsumtionen i olika europeiska länder (inklusive Sverige) kan man få på webbplatsen för European Centre for Disease Prevention and Control när man klickar på "Health Topics" och sedan väljer "Health topics A-Z" och därefter "Antimicrobial resistance". Där kan man sedan välja t.ex. "Antimicrobial resistance interactive database (EARS-Net)" under Interactive Databases. En annan viktig källa till information om antibiotikakänslighet i form av MIC-värden är webbplatsen EUCAST (EUropean Committee on Antimicrobial Susceptibility Testing), där man kan söka i en databas på ett specifikt antibiotikum eller en viss bakterie.
Se även Resistensundersökningar här på VetBact.
Uppdaterad: 2025-06-25.
Ett aseptiskt arbetssätt innebär att man förhindrar att sjukdomsframkallande mikroorganismer kontaminerar det material man arbetar med, utan att man använder kemiska desinfektionsmedel. Kemiska desinfektionsmedel (= germicider) är substanser, som används för att hämma tillväxten av eller eller döda mikroorganismer på antingen föremål eller på en kroppsyta. Aseptik avser alltså att förhindra uppkomsten av mikroorganismer till skillnad mot antiseptik som avser att bekämpa redan befintliga mikroorganismer med hjälp av antiseptiska medel.
Ett antiseptikum (antiseptiskt medel) är en substans, som används för att hämma tillväxten av eller eller döda mikroorganismer på en kroppsyta.
En baktericid är en substans, som dödar bakterier. Ett bakterostatikum är en substans, som hämmar tillväxt av bakterier.
En germicid är en substans, som dödar inte bara bakterier utan även andra mikroorganismer.
Uppdaterad: 2014-01-22.
.")
Fig. 1. Vid ytspridning fördelas 0,1 ml med en steril rackla på den solida agarn i detta fall mCCDA [Bild: Moa Skarin & Ingrid Hansson (BVF, SLU)]. - Klicka på bilden för att förstora den.
.")
Fig. 2. Djupspridning. I detta fall djupsprids i VRGG för beräkning av antalet bakterier tillhörande familjen Enterobacteriaceae. [Bild: Tomasz Dzieciolowski, Ingrid Hansson & Karl-Erik Johansson (BVF, SLU)] - Klicka på bilden för att förstora den.
Bakterier kan räknas på olika sätt och här följer tre principiellt olika metoder:
Bestämning av CFU (Colony Forming Unit). Prov som ska analyseras späds och sprids på/i en agarplatta, som inkuberas under olika betingelser beroende på analysmetod, dvs vilka bakterier frågeställningen gäller. Metoden är relativt noggrann under förutsättning att spädningen är gjord på ett korrekt sätt och att bakterierna inte bildar aggregat i suspensionskulturer. Nackdel: svaret erhålles först efter minst ett dygn beroende på vilken agar som används och vilken bakterie man letar efter. Se även Kolonibildande enhet.
Inom livsmedelsmikrobiologi används ytspridning eller djupspridning beroende på vilken analysparameter det gäller. Från en tiofaldig spädningsserie används 0,1 ml och 1,0 ml av de olika spädningsstegen för ytspridning resp. djupspridning. Vid ytspridning racklas 0,1 ml från respektive spädning över hela agarytan (se Fig. 1). Medan vid djupspridning blandas 1,0 ml från spädningssteget med den "smälta" agarn direkt i petriskålen (se Fig. 2). Bakterierna blir då fördelade i hela volymen av agar och inte bara på ytan. Den smälta agarn agarn har temperaturen 45° C, vilket inte dödar bakterierna, men håller blandningen flytande.
Spektrofotometrisk bestämning av ljusspridning. Metoden ger ett relativt mått på antalet bakterier, men kan kalibreras med hjälp av en standardkurva. Se Tillväxtkurva.
Uppdaterad: 2020-02-13.
Uppdaterad: 2013-03-03.
.")
Figuren visar en schematisk bild av hur en biofilm bildas. Två olika bakteriearter (A och B) lever först planktoniskt. En av bakterierna (1), som har vidhäfningsförmåga fäster till ytan och börjar där att dela sig (2). Efter en tid har den bildat ett monolager och polysackarder (glykokalyx) börjar utsöndras (3). Så småningom börjar monolagret att växa på höjden och en koloni, som är inkapslad i glykokalix bildas (4). Detta är början till en biofilm, som kan innehålla flera olika arter av bakterier och även svampar (av typ jäst eller mögel). När biofilmen har vuxit till en viss storlek, lossnar bakterier (6) eller klumpar av bakterier, som kan fästa till nya platser och bilda mer biofilm.
Bild: Karl-Erik Johansson (BVF, SLU) - Klicka på bilden för att förstora den.
Bakterier som växer fritt i flytande medium sägs vara planktoniska, men vissa bakterier kan övergå till att växa på fasta substrat och bildar då s.k. biofilmer. Biofilmer kan finnas på levande eller döda ytor och förekommer i naturen, på material vid industriella processer och inom sjukvården. Biofilmer utgörs av populationer (en eller flera) av bakterier, som fäster till en yta och till varandra samt är inneslutna i ett nätverk (matris) av biopolymerer. Bildning av biofilm startar med att bakterier fäster på en yta med hjälp av t.ex fimbrier (pili) där de binder irreversibelt och först växer som ett monolager. Därefter bildar de flera lager och producerar någon form av biopolymer (extracellulär matris), som ofta består av samma material som kapseln, men i en lösare struktur. Biopolymeren består alltså av polysackarider och kallas glykokalyx (som kapseln). Dextran är ett exempel på en sådan polysackarid. Biofilm består av en eller flera bakteriepopulationer (arter), glykokalyx, DNA och proteiner.
En bakterieart, som inte själv kan fästa vid ytor, kan ofta fastna på andra bakteriers glykokalyx och växa i form av en biofilm. Bakterier i biofilmer är mer motståndskraftiga mot antibiotika, rengöringsmedel och fagocytos än vad planktoniska bakterier är. Bakterier (Streptococcus spp.) kan växa i form av biofilmer på tänder (= plaques), på implantat (t.ex hjärtklaffar), i plastslangar som transporterar näringsämnen mm. Hos patienter med cystisk fibros kan Pseudomonas aeruginosa växa i form av biofilm i lungorna. I en biofilm kan bakterier kommunicera med varandra genom att skicka ut kemiska signalsubstanser (quorum sensing) för att reglera genuttrycket i hela populationen.
Länk till videomontage på YouTube som rekommenderas:
Biofilm: "What Are Bacterial Biofilms?"
Quorum sensing: "Quorum sensing, bacteria talks"
Uppdaterad: 2021-01-28.
.")
Schematisk bild av cellhöljet hos grampositiva och gramnegativa bakterier. NAG-NAM står för N-acetylglukosamin och N-acetylmuraminsyra, som är cellväggens byggstenar. TA och LTA står för teikonsyra resp. lipoteikonsyra, som stabiliserar cellväggen och förankrar den i plasmamembranet hos grampositiva bakterier. LPS står för lipopolysackarid och BLP står för Browns lipoprotein, som binder cellväggen till yttermembranet i gramnegativa bakterier.
Bild: Karl-Erik Johansson (BVF, SLU). - Klicka på bilden för att förstora den.
Cellvägg och cellmembran är mycket viktiga strukturer hos bakterier eftersom de utgör bakteriens kontaktyta med omgivningen. För bakterier brukar man tillsammans kalla cellvägg och cellmembran samt eventuellt yttermembran för cellhölje. Skillnader i strukturer hos cellhöljet kan användas för differentiering av olika grupper av bakterier t.ex. genom gramfärgning. För vissa antibiotika utnyttjas det faktum att det är stora skillnader mellan mellan strukturer i bakteriers cellhölje och cellhöljet hos högre organismer (eukaryota celler). Det finns två olika huvudtyper av strukturer hos bakteriernas cellhöljen och eftersom det kan utnyttjas vid gramfärgning, så kallas dessa bakterier för grampositiva eller gramnegativa. Skillnaderna mellan grampositiva och gramnegativa cellhöljen visas mycket förenklat i figuren.
Hos grampositva bakterier är cellväggen 20-80 nm tjock medan hos gramnegativa bakterier är den bara 5-10 nm tjock. Cellväggen består av peptidoglykan, som är uppbyggd av repeterande enheter av N-acetylglukosamin (NAG) och N-acetylmuraminsyra (NAM). Peptidoglykankedjorna är tvärbundna med varandra genom peptidbryggor och bildar ett kontinuerligt nätverk runt bakterien och kan betraktas som en jättemolekyl. Gramnegativa bakterier har också ett yttermembran innehållande lipopolysackarid (LPS) som skydd mot omgivningen. Både plasmamembran och yttermembran är ca 8 nm tjocka och innehåller proteiner som är viktiga för bakteriens alla funktioner. Vissa författare innefattar både peptidoglykan och yttermembran hos gramnegativa bakterier i begreppet cellvägg. Periplasman är det utrymme, som finns mellan cellvägg och plasmamembran i gramnegativa bakterier. Vissa författare definierer periplasman, som det utrymme, som finns mellan plasma- och yttermembran. Periplasman är geléartad och innehåller enzymer, som är viktiga för bakteriens metabolism.
Förutom ovanstående komponenter kan bakterier ha flageller, fimbrier och kapsel på cellytan (visas ej i figuren).
Cellhöljet är naturligtvis livsnödvändigt för bakterien eftersom dess omgivning kan vara väldigt ogästvänlig och höljet utgör alltså ett skydd för bakterien. Det finns olika enzymsystem, som är bundna till plasmamembranet, t.ex. de som ingår i elektrontransportkedjan och en del av de enzymer, som deltar i syntesen av cellväggen. Den molekylära motor, som består av olika proteinsubenheter och driver flagellers rörelse är förankrad i bakteriens plasma membran. Många bakterier har s.k. sekretionssystem, som är av stor betydelse för deras sjukdomsframkallande förmåga. Sekretionssystemen består också av ett antal olika membranbundna proteinsubenheter. Poriner är membranbundna proteiner som kan transportera sustanser genom plasmamembranet och yttermembranet.
Det finns bakterier, som har en helt annan uppbyggnad av cellhöljet än vad som beskrivs i figurerna ovan.
Fylum Mycoplasmatota
Bakterier som tillhör fylumet Mycoplasmatota, som omfattar de organismer, som i vanligt tal kallas mykoplasmer, saknar cellvägg eftersom de inte kan tillverka peptidoglykanen. Dessa bakterier lever oftast i fysiologiska miljöer där det mekaniska skyddet av cellväggen mot osmotisk lysering inte behövs. Mer information om mykoplasmer finns i en föreläsningsserie.
Familjen Chlamydiaceae
Man trodde länge att bakterier inom familjen Chlamydiaceae inte har någon peptidoglykan, utan istället cysteinrika proteiner, som stabiliserar yttermembranet, genom disulfid-bindningar. För några år sedan visade dock en forskargrupp att retikulärkropparna hos klamydier har en tunn peptidoglykan, som är av betydelse vid celldelningen, vilken hos klamydier sker i inklusioner hos värdcellen. I föreläsningen om klamydia sägs det att klamydia saknar peptidoglykan, vilket alltså inte är korrekt med dagens kunskap.
Ordning Corynebacteriales
Ordning Corynebacteriales dit familjerna Corynebacteriaceae, Mycobacteriaceae och Nocardiaceae hör, har ett cellhölje, som mest liknar det vilket grampositiva bakterier har. Desssa bakterier har alltså ett plasmamembran, peptidoglykan och utanför peptidoglykanen ett polysackaridlager (arabinogalaktan) samt mykolsyra, som med sina kolvätekedjor binder till ett enkellager av lipider. Mykobakterier sägs vara syrafasta eftersom de har ett ovanligt stabilt cellhölje.
Uppdaterad: 2024-03-20.
Uppdaterad: 2013-02-23.
.")
Figuren visar en schematisk bild av elektrontransortkedjan i bakterier med aerob metabolism. Vätejoner (protoner) pumpas ut (blå pilar) ut cellen med hjälp av tre olika enzymkomplex och det bildas en protongradient över plasmamembranet. Protongradienten utgör den energi (protonpotential), som används av ATP-syntaset för att producera ATP. Elektronerna hoppar stegvis mellan de olika komponenterna i enzymkomplexen, som har ökad elektronegativitet från vänster till höger i figuren. Ju högre elektronegativitet desto starkare är attraktionen av elektroner. Syre är den komponent i systemet, som har den högste elektronegativiten och det är också den slutliga elektronacceptorn.
Bild: Karl-Erik Johansson (BVF, SLU) - Klicka på bilden för att förstora den.
Under glykolysen bildas två molekyler pyruvat, som har 3 kolatomer vardera, från en glukosmolekyl, som har 6 kolatomer. Under aeroba betingelser och om den aktuell bakterien har aerob metabolism, så fermenters inte pyruvatmolekylen, utan kan istället omvandlas till acetyl-Coenzym A (acteyl-CoA). Acetylgruppen i acetyl-CoA har två kolatomer eftersom koldioxid (CO2) har spjälkats av och kan nu gå in i Krebscykeln (= citronsyracykeln), som en acetylgrupp. Krebscykeln beskrivs i ett separat avsnitt och är ett försteg till elektrontransportkedjan (= andningskedjan).
Alla enzymer, som krävs för bakteriers elektrontransportkedja är membranbundna liksom i eukaryota celler, men hos bakterier finns dessa molekyler i plasmamembranet eftersom bakterier inte har några mitokondrier. Vätejongradienten, som driver ATP-syntes, genereras alltså över plasmamembranet. Elektrontransportkedjan består av en serie enzymkomplex, som stegvis tar hand om de elektroner, som bildas när NADH och FADH2 (från Krebscykeln) oxideras till NAD+ resp. FAD. Samtidigt pumpas vätejoner (protoner) ut ur bakteriecellen. När dessa vätejoner sedan passerar att annat membranbundet enzymkomplex (ATP-syntas) på sin väg tillbaka in i cytoplasman, så genereras ATP samtidigt som elektronerna slutligen tas om hand av syre och vatten bildas med de vätejoner, som nu finns i cytoplasman.
Det finns olika varianter av elektrontransportkedjan och Escherichia coli, till exempel, saknar cytokrom C-oxidas liksom de flesta andra bakterier inom familjen Enterobacteriaceae. Dessa bakterier har istället ett terminalt cytokrom bo3 oxidas. Vissa bakterier har enzymsystem för anaerob respiration, där andra molekyler än syre utgör den terminala elektronacceptorn. Exempel på sådana molekyler är: nitrat, nitrit, trevärt järn (Fe3+), sulfat, koldioxid, och små organiska molekyler som fumarat. Vid anaerob respiration talar man om terminala reduktaser i stället för terminala oxidaser.
Uppdaterad: 2021-10-06.
Med enkelfärgningsmetoder kan man avgöra om ett prov innehåller stavar eller kocker, men ej om de är grampositiva eller gramnegativa. För att avgöra om bakterierna är grampositiva eller gramnegativa kan man använda Gramfärgning, som är en differentialfärgningsmetod.
Uppdaterad: 2013-02-24.
.")
Honungsbin i en bikupa i Västeråker. Detta bisamhälle har dock inte drabbats av amerikansk yngelröta.
Foto: Karl-Erik Johansson (BVF, SLU).
Lytiska bakteriofager (fager) kan användas för att döda bakterier och, i princip, också för att behandla bakteriella infektionssjukdomar. Eftersom allt fler bakterier blir resistenta mot de antibiotika, som kan användas för behandling, så har intresset för användning av fagterapi ökat. Det finns dock både problem och fördelar med att använda fagterapi för behandling. En fördel är att fager är oerhört specifika och inte påverkar andra bakteriearter än sin egen värdart och givetvis inte heller eukaryota celler. Detta innebär t.ex. att värddjuret tarmflora inte alls påverkas av behandling med fager. Den höga specificiteten är dock även en nackdel eftersom fagen är specifik inte bara mot en viss art, utan även mot enbart en viss stam av denna art. Detta betyder att för en lyckad behandling krävs inte bara en typ av fager, utan en hel cocktail av fager, som är specifika med de patogena stammar, som kan tänkas finnas av arten i fråga. Metoden är fortfarande på försöksstadiet, men många forskare anser att den har stor potential både inom human- och veterinärmedicin eftersom resultat av vissa lyckade experiment har publicerats. Man har t.ex. framgångsrikt använt fagterapi för att behandla amerikansk yngelröta hos honungsbin, som orsakas av Paenibacillus larvae.
Uppdaterad: 2018-05-30.
Bakterier kan ha olika typer av utskott från cellytan, som kallas flageller och fimbrier. Fimbrier kallas även pili.
Flagellens främsta uppgift är att ge bakterierna rörlighet. De kan alltså "simma" med hjälp av sina flageller. Rörlighet kan vara viktig för bakteriers möjlighet att orsaka sjukdom och därför brukar flageller räknas som en patogenicitetsfaktor. Flageller är vanligen 10-30 nm i diameter och 5-15 µm långa. Bakterier kan indelas efter antalet flageller och hur de är arrangerade på cellytan enligt följande:
Flagellen är uppbyggd av tre delar, som består av olika proteiner:
Bakterier inom fylum Spirochaetota har s.k. periplasmatiska flageller (= axialfilament = endoflageller), som är lokaliserade till det periplasmatiska utrymmet och ger bakterierna en mycket karaktäristisk korkskuvsliknande rörelse.
Fimbrier (= pili) är en annan typ av hårlika utskott, som i vissa undantagsfall (typ IV pili) kan ge bakterier rörlighet (t.ex. hos Psueudomonas sp.), men framför allt bidrar till bakteriers vidhäftningsförmåga (= adhesionsförmåga). S.k. F-pili eller sex-pili ger bakterier möjlighet att utbyta genetiskt material (DNA).
Uppdaterad: 2025-05-23.
Se Taxonomi och Fylogeni inom bakteriologi.
Uppdaterad: 2023-04-12.
Med termen genom menar man den kompletta uppsättningen av genetisk material i en cell. Genomstorlek brukar uttryckas i Mb eller Mbp (= megabaspar). Alltså är 1 Mbp = 1 000 000 bp. Storleken på bakteriegenom varierar mellan 0,5 och 10 Mbp. Bakteriegenom består av kromosom(er) och eventuellt även plasmid(er). Bakterier är haploida till skillnad från högre organismer, som är diploida. Könsceller från högre organismer är emellertid också haploida. Den fullständiga genomsekvensen har bestämts för ca 30 000 bakteriestammar och det finns ytterligare ca 20 000 publicerade genomprojekt för bakterier, där man inte har kombinerat delsekvenserna till en enda fullständig sekvens (en s.k. kontig).
Kromosomen utgör primärt genetiskt material, som är helt nödvändigt för bakterien. Bakterier har i allmänhet en cirkulär kromosom, men det finns undantag (se nedan).
Plasmider (see också Genom ovan) utgör sekundärt genetiskt material, som inte alltid är livsnödvändigt för bakterien. Bakterier, som bär på plasmider, har i allmänhet en eller flera cirkulära plasmider men det finns också undantag. Plasmider utgör i allmänhet upp till 10% av genomet och de replikeras oberoende av kromosomen.
Uppdaterad: 2020-09-23.
.")
Figuren visar en mycket förenklad bild av bakteriell glykolys och fermentation där slutprodukten är laktat (mjölksyrans joniserade form). Nettovinsten vid glykolysen är 2 ATP och 2 NADH för varje glukosmolekyl. De två molekylerna NADH åtgår dock vid fermentationen av pyruvat till laktat. Pyruvat fungerar som en elektronacceptor och kan oxidera NADH till NAD+, som sedan återanvänds för att oxidera glukos igen.
Bild: Karl-Erik Johansson (BVF, SLU) - Klicka på bilden för att förstora den.
Alla bakterier av betydelse inom veterinär- och humanmedicin är kemoorganoheterotrofa, vilket betyder att de får sin energi från organiska kolföreningar, som också utnyttjas som kolkälla. Enda undantaget är cyanobakterier, som är fotolitoautotrofa, d.v.s. de får sin energi från ljus och de utnyttjar oorganiskt kol i form av CO2 (koldioxid) som kolkälla.
Bakterier kan alltså utvinna energi genom oxidation av kolhydrater (framför allt glukos) och dessa bakterier sägs vara sackarolytiska. En del andra bakterier kan istället använda aminosyror eller lipider som energikälla, men detta är ovanligare och dessa bakterier sägs vara asackarolytiska. Kolhydrater ingår också i en mängd olika cellulära processer hos bakterier och bakteriell kolhydratmetabolism är mycket diversifierad. Detta är förklaringen till att bakterier kan växa och frodas i praktiskt tagat alla miljöer på vår planet.
Glukos, som är en monosackarid med 6 kolatomer (hexosmonosackarid), är en av de viktigaste kolhydratmolekylerna, som bakterier kan utvinna energi ifrån. Detta sker genom en oxidationsprocess, som kallas glykolys och vid glykolysen bryts glukos ner till pyruvat. Bakterier kan också bryta ner bl.a. laktos och mannitol till pyruvat genom glykolys. Laktos är en disackarid, som består av två hexosmonosackarider (galaktos och glukos). Mannitol är en hexosmonosackaridalkohol. Om en bakterie har de rätta enzymsystemen så kan även dessa kolhydrater oxideras genom glykolys.
Under anaeroba förhållanden kan pyruvat omvandlas till en rad olika produkter, beroende av vilka enzymsystem mikroorganismen ifråga har. Denna process kallas fermentation och genom att undersöka vilka kolhydrater en viss bakterie kan fermentera och vilka slutprodukter, som bildas, kan man identifiera bakterien ifråga.
Vissa bakterier kan också omvandla aminosyran tryptofan till pyruvat om de har enzymet typtofanas (kan påvisas genom indoltestet). Puruvat kan sedan ta olika vägar i metabolismen, beroende på om O2 (syrgas) finns tillgängligt och beroende på vilka enzymsystem, som bakterien har. Om O2 saknas och om bakterien är fakultativt anaerob, syretolerant eller anaerob kan puruvat omvandlas vidare genom fermentation.
Fermentation är en metabolisk process, som inte kräver O2 och som kan utnyttjas av väldigt många olika bakterier. Fermentationen i kombination med glykolysen resulterar i produktion av ATP, som är den viktigaste energiformen för bakterier och andra organismer. Exempel på slutprodukter efter fermentationen är: mjölksyra (laktobaciller), etanol (jäst), 2,3-butandiol (Klebsiella spp.), myrsyra (Shigella spp.), smörsyra (Clostridium sp.) och blandade syror (E. coli m.fl.). Glykolys i kombination med fermentation är inte någon effektiv process för att utvinna energi eftersom kolhydraten inte oxideras fullständigt till CO2 och H2O. Därför bildas det bara 2 ATP-molekyler för varje kolhydratmolekyl som oxideras och fakultativa anaerober växer alltid bättre i närvaro av O2.
I avsnitten om Krebscykeln och Elektrontransportkedjan beskrivs hur den kemiska energin i pyruvat kan utvinnas på ett effektivare sätt.
Uppdaterad: 2019-10-10.
Gramfärgning är en s.k. differentialfärgningsteknik eftersom man kan skilja på två stora grupper av bakterier genom denna metod. Dessa två grupper är grampositiva och gramnegativa bakterier, som färgas mörklila respektive skära till röda.
Grampositiva bakterier har en tjock cellvägg (peptidoglykan), som består av flera lager och kan liknas vid ett nätverk. Gramnegativa bakterier har en mycket tunnare cellvägg och dessutom ett yttermembran. Kristalviolett (KV+), som är det primära färgämnet, binder till negativt laddade grupper på bakterien och färgar den lila. Man använder sedan jod (I-), som bildar ett stort komplex (KV-I) med KV och därmed binds färgen till bakterien. När grampositiva bakterier behandlas med avfärgningsvätskan (etanol-aceton), så dehydreras bakterien och färgen stannar kvar. När gramnegativa bakterier behandlas med avfärgningsvätskan, så löses yttermembranet upp och peptidoglykanen exponeras, så att även KV-I-komplexet tvättas ut. Därefter gör man en motfärgning med safranin eller basiskt fuksin, som gör att gramnegativa bakterier blir skära eller röda.
Medlemmar av fyla Bacillota och Actinomycetota (undantag: genus Mycobacterium).
Medlemmar av fyla Bacteroidota, Campylobacterota, Cyanobacteria, Fusobacteriota, Pseudomonadota (undantag: vissa medlemmar av ordningen: Rickettsiales), Spirochaetota och Thermodesulfobacteriota.
Uppdaterad: 2025-05-23.
.")
Fig. 1. Bilden visar kolonier av några olika bakteriearter, som uppvisar olika hemolysmönster. Vid fotograferingen har kolonierna dels belysts ovanifrån. Normalt är det allra lättast att observera hemolys med belysning underifrån och genom att titta på plattan i "rätt" vinkel. Följande bakterier har använts för att illustrera hemolys:
A. Streptococcus uberis, som inte ger någon hemolys. Detta brukar ibland lite olyckligt kallas för γ-hemolys.
B. Streptococcus agalactiae, som ger en klar (fullständig) β-hemolys.
C. Streptococcus dysgalactiae (underart ej definierad), som ger ofullständig grönaktig α-hemolys.
D. Staphylococcus pseudintermedius, som ger dubbelhemolys.
- Klicka på bilden för att förstora den.
.")
Fig. 2. Bilden visar samma agarplattor som i Fig. 1, men vid fotograferingen har kolonierna nu belysts underifrån då det är allra lättast att observera hemolysen. - Klicka på bilden för att förstora den.
Hemolys innebär att röda blodkroppar (erytrocyter) spricker sönder (hemolyserar) och släpper ut cellinnehållet (hemoglobin). Vissa bakterier producerar s.k. hemolysiner, som ger dem hemolytisk förmåga. De flesta hemolysiner är proteiner (enzymer eller poriner), men det finns även andra typer av hemolysiner som rhamnolipider och biologiska detergenter.
Hemolysiner är membranförstörande exotoxiner och de kan indelas i två grupper: toxiner med enzymatisk aktivitet och kanalbildande toxiner (= poriner).
Poriner är uppbyggda av subenheter, men utsöndras av bakterien i monomerisk form. I värddjurets cellmembran aggregerar monomererna till kanalbildande polymerer (heptamerer), som gör att jongradienten över värdcellens plasmamembran inte kan upprätthållas och det osmotiska trycket i cellen ökar tills den lyserar.
En funktion hos hemolysiner är att bakterien kan utnyttja hemolys för att frigöra och utnyttja näringsämnen från värddjurets celler. Järn t.ex., är essentiellt för många patogena bakterier, men finns bara närvarande i mycket låga koncentrationer utanför celler. Om bakterien har tillgång till fritt hemoglobin så kan den utnyttja det järn, som är bundet till hemgrupperna i hemoglobinet. Hemolysiner verkar inte bara på erytrocyter utan kan också lysera andra typer av celler.
Genom odling på blodagar kan bakterier differentieras, på basis av deras förmåga att producera hemolysiner. Hemolysen kommer att ge upphov till en uppklarningszon av blodagarn runt kolonin. Bakterier kan orsaka olika typer av hemolys:
Notera att stafylokockernas α-hemolysin orsakar fullständig hemolys (β-hemolys) medan deras β-hemolysin orsakar ofullständig hemolys (α-hemolys).
Den hemolysinproducerande förmågan kan variera mellan olika stammar av en viss bakterieart. Blodagarplattor med bakterier, som uppvisar olika hemolysmönster visas i Fig. 1 och 2. Notera att i Fig. 1A och 2A kan man inte se någon hemolys över huvud taget, men observera att det finns stammar av S. uberis, som ger α-hemolys. I Fig. 1B kan man ana en tunn hemolyszon och i Fig. 2B syns den klara β-hemolysen tydligt runt alla kolonier. I Fig. 1C kan man ana hemolys runt vissa kolonier och i Fig. 2C kan man tydligt se den gröna α-hemolysen runt vissa kolonier (vita pilar). I Fig. 1D kan man se den yttre hemolyszonen (vit pil) och i Fig. 2D kan man se både den inre klara β-hemolyszonen och den yttre grumliga α-hemolyszonen (vita pilar).
Notera också att alla kolonier av Streptococcus dysgalactiae inte ger upphov till α-hemolys trots att den använda stammen är ren med avseende på art. Det kan dock vara så att olika stammar (eller kloner) av en och samma art uppvisar olika hemolysmönster.
Uppdaterad: 2025-05-23.
 För att kunna odla ut bakterier från vissa typer av prover (bl.a. livsmedel och foder) är det viktigt att finfördela provet i spädningssbuljong utan att skada bakterierna. En Stomacher® är en apparat, som används för homogenisering av prover inom bl.a. livsmedelsmikrobiologi. En stor mängd viabla mikroorganismer kan frigöras från olika typer av livsmedelsprover med denna metod. Apparaten behöver inte steriliseras eftersom proven läggs i sterila plastpåsar till vilka sterilt medium tillsätts.
För att kunna odla ut bakterier från vissa typer av prover (bl.a. livsmedel och foder) är det viktigt att finfördela provet i spädningssbuljong utan att skada bakterierna. En Stomacher® är en apparat, som används för homogenisering av prover inom bl.a. livsmedelsmikrobiologi. En stor mängd viabla mikroorganismer kan frigöras från olika typer av livsmedelsprover med denna metod. Apparaten behöver inte steriliseras eftersom proven läggs i sterila plastpåsar till vilka sterilt medium tillsätts.
A. Homogenisatorn, som laddas med en plastpåse innehållande provmaterial och odlingsmedium
B. Ställ för påsarna under inkubering i värmeskåp
C. Ställ, som används vid invägning av undersökningsmaterialet och tillsättning av odlingsmedium
D. Sterila plastpåsar för homogenisering och inkubering
Klicka på bilden om du vill förstora den.
Uppdaterad: 2025-05-23.
Infektion och inflammation är två begrepp som ofta kopplas ihop, men trots detta är de inte synonyma.
Infektion innebär att vävnader i kroppen har koloniserats av mikroorganismer (bakterier, svamp, virus eller protozoer), som fortsätter att föröka sig och orsaka skada på vävnader. Om mikroorganismen i fråga inte orsakar skada utan tillhör normalfloran och kanske t.o.m. är nyttig för sitt värddjur pratar man istället om kolonisation.
Inflammation är kroppens försvarsreaktion på ett angrepp av mikroorganismer eller en skada (t.ex, slag, värme, kyla eller allergi). Vissa sjukdomar kan också orsaka inflammation. Kroppens försvarsreaktionerna karaktäriseras bl.a. av att de perifera blodkärlen vidgas för att bli permeabla för blodplasma och immunceller (vita blodkroppar), som då kan lämna blodomloppet och söka upp och oskadliggöra mikroorganismer eller reparera skador på kroppens celler.
När det gäller ektoparasiter ser man ibland begreppet infestation i stället för infektion och framför allt i amerikansk litteratur. Detta uttryck bör dock undvikas eftersom det kan skapa förvirring.
Uppdaterad: 2019-03-04.
Varför är järn essentiellt?
Järn är essentiellt för nästan alla livsformer, därför att det är en nödvändig komponent för energiproduktion, DNA-replikation, transport av syre och skydd mot oxidativ stress. Bakterier utgör i det avseendet inte något undantag och patogena bakterier måste kunna ta upp järn i värddjuret för att kunna föröka sig och orsaka sjukdom.
Hur kan värddjur utnyttja att järn är essentiellt för bakterier?
En försvarsmekanism, som högre organismer har mot bakterier är att minimera tillgången på fritt järn. Huvuddelen av det järn som finns hos ryggradsdjur är intracellulärt och bundet till det järnlagrande proteinet ferritin eller komplexbundet till porfyrinringen hem som utgör en kofaktor till hemoglobin och myoglobin. Dessutom finns det ett järntransporterande protein i serum, som kallas för transferrin och som binder järn extremt kraftigt.
Hur kan bakterier överlista värddjuret?
Det skulle inte var möjligt för bakterier att överleva i sina värddjur om de inte hade utvecklat speciella mekanismer för att kunna utvinna det bundna järnet. De allra flesta patogena bakterier har järnupptagningsmekanismer, som har så hög affinitet för järn att de kan konkurrera med värddjurets järnbindande substanser. Dessa mekanismer kan vara baserade på sideroforer, hem-utvinning eller transferrin/laktoferrin-receptorer.
Borrelia burgdorferi, som orsakar borrelios, utgör ett undantag från ovanstående regel eftersom denna bakterie utnyttjar mangan istället för järn. D.v.s. de enzymer, som normalt innehåller järn, är modifierade och innehåller i B. burgdorferi mangan. Medlemmar av släktet Lactobacillus anses också kunna överleva utan tillgång på järn, men detta har ifrågasatts i nya studier.
Uppdaterad: 2017-11-10.
Många bakterier har en s.k. kapsel, som oftast består av polysackarider och ibland även av andra substanser. Kapseln utgör en struktur utanför bakteriernas cellhölje och man kan också säga att kapseln är det yttre höljet av bakterien. Kapseln är ett väldefinierat lager, som kan utgöra en virulensfaktor hos både grampositiva och gramnegativa bakterier. Om den amorfa och viskösa sekretionen, som utgör kapseln, diffunderar ut i omgivningen och återstår som att löst odefinierat lager utan tydlig yttre gräns, brukar men istället tala om ett slemlager. Kapsel och slemlager brukar sammanfattningsvis kallas för glykokalyx. En biofilm bildas när många bakterier innesluts i ett gemensamt slemlager.
En bakteries polysackaridkapsel kan bestå av väldigt många olika linjära polysackarider. Dessa polysackarider är uppbyggda av repeterande subenheter, som består av en till sex monosackarider. Diversiteten är enorm och man har visat att bara Escherichia coli kan producera ca 200 olika typer av polysackarider. Kapseln kan också bestå av peptidoglykan (som cellväggen), poly-muraminsyra (en av cellväggens komponenter), glykoproteiner och poly-D-glutaminsyra som hos Bacillus anthracis. För att kunna se kapseln i ett mikroskop, kan man använda en speciell kapselfärgningsmetod med metylenblått och detta har ytnyttjats för B. anthracis (se bakteriesidan).
Kapseln betraktas som en patogenicitetsfaktor, eftersom den kan öka bakteriers förmåga att orsaka sjukdom. Kapseln skyddar bakterien från fagocytos av makrofager. Kapseln innehåller vatten och skyddar därför bakterier mot uttorkning. Kapseln ökar bakteriers förmåga att binda till ytor och till varandra. När bakterier bildar en biofilm, är de mindra känsliga för antibiotika och vissa andra bakteriostatiska ämnen. Kapseln kan också skydda bakterier mot angrepp av vissa bakteriofager.
Uppdaterad: 2020-04-09.
Robert Koch var en tysk läkare (1843-1910), som intresserade sig för sambandet mellan mikroorganism och sjukdom. Koch formulerade fyra kriterier (Kochs postulat), som måste vara uppfyllda för att bevisa att en viss mikroorganism har orsakat en viss sjukdom (för virus gäller dock andra kriterier).
Kochs postulat
I orginalversionerna av postulaten gällde ska, men med dagens kunskap måste man använda bör eftersom det finns många undantag. Det första kriteriet var man tvungen att överge, när det upptäcktes att det finns symptomlösa (asymptomatiska) bärare av vissa mikroorganismer (subkliniska infektioner). Det andra kriteriet måste ibland också överges eftersom det finns mikroorganismer, som inte går att odla. Det tredje kriteriet är inte heller alltid giltigt eftersom yttre omständigheter kan påverka resultatet av en experimentell infektion.
Om alla kriterier är uppfyllda har man bevisat samband mellan mikroorganism och sjukdom, men om inte alla kriterier är uppfyllda, så kan det ändå finnas ett samband.
År 1996 publicerades ett antal nya kriterier för att bevisa ett samband mellan mikroorganism och sjukdom (Fredricks and Relman, Clin. Microbiol. Rev. 9:18-33). Dessa kriterier är baserade på detektion av specifika DNA-sekvenser i samband med sjukdom.
Uppdaterad: 2017-09-27.
Kolhydrater är en mycket viktig energikälla för bakterier och namnet kolhydrat syftar på att det är substanser, som innehåller kol (C), samt väte (H) och syre (O) i samma proportioner som i vatten (några undantag finns dock). Bakterier kan dock inte utnyttja di-, tri- eller polysackarider för glykolys om de inte har enzymsystem, som kan bryta ner dessa till monosackarider. Exempel på kolhydrater, som bakterier kan metabolisera genom fermentation under bildning av en syra, ges nedan.
Fermentationen kan detekteras genom tillsätta en pH-indikator till odlingsmediet (flytande eller fast) och detta kan alltså användas för differentiering av bakterier.Olika laboratorier använder olika kombinationer av kolhydrater i flytande media och beroende på vilken bakterie man misstänker, så görs valet av kombination. Nedan listas kolhydrater, som kan användas för identifiering av bakterier. Ibland vill man utföra jäsningstesterna på agarplattor eftersom det i vissa fall ger säkrare resultat.
Uppdaterad: 2019-02-24.
Koliforma bakterier används i livsmedelsproduktion som indikatorbakterier för allmän hygien. Vid mikrobiologisk analys av vatten används koliformer alltid som indikatorbakterier.
Exempel på koliformer: Escherichia spp. (inkl. E. coli), Citrobacter spp. Enterobacter spp., Hafnia spp. och Klebsiella spp.
Uppdaterad: 2015-06-17.
.")
Kolonibildande enheter (CFU) kan kvantifieras med hjälp av djupspridning av bakterier tillhörande familjen Enterobactericaeae. I detta fall VRGG från ett livsmedelsprov.- Klicka på bilden för att förstora den.
.")
Kolonibildande enheter (CFU) från ett urinprov kan kvantifieras genom att en bestämd mängd urin fördelas på en CLED agar. I detta fall växer det >100 000 CFU/ml. - Klicka på bilden för att förstora den.
Mängden bakterier i ett fast prov, t ex livsmedel, uttrycks ofta som kolonibildande enheter (colony forming units) per gram, (CFU/g), medan antalet bakterier i ett flytande prov uttrycks som kolonibildande enheter per ml (CFU/ml). Detta värde kan bestämmas genom att man först gör en tiofaldig spädningsserie av provet. Därefter tas en känd volym (oftast 1 ml eller 0,1 ml) från varje spädning och appliceras på lämpliga agarplattor. Efter inkubering räknas antalet kolonier på den agarplatta, som har 15-150 kolonier, vilket anses ge det mest noggranna resultatet. Vid beräkningen av det faktiska antalet bakterier i provet skall spädningsfaktorn också inkluderas.
Vid bedömning av urinprov är antalet bakterier per ml (CFU/ml) av stor betydelse. Urin tas med en ögla med en känd volym (1 µl) och fördelas på en agarplatta så att räkning av antal kolonier underlättas. Mängden bakterier i urin brukar klassificeras enligt följande:
Observera att detta sätt att bestämma mängden bakterier i urin endast ger en grov uppskattning av det faktiska antalet.
Bakteriuri definieras som närvaron av bakterier i urin i nästan renkultur och med en koncentration av mer än 100 000 CFU/ml. En patient sägs ha urinvägsinfektion (UVI) när det finns bakteriuri och kliniska symtom på infektion.
Anledningen till att man använder begreppet CFU är att det inte är säkert att varje koloni härstammar från en enda bakterie eftersom vissa bakteriearter lätt aggregerar i suspensionskulturer. Endast levande (egentligen odlingsbara) bakterier kommer att ge upphov till kolonier.
Uppdaterad: 2020-02-20.
.")
Förenklad bild av Krebscykeln där man ser hur pyruvat omvandlas till acetyl-CoA, som reagerar med oxaloacetat och bildar citrat (därav det alternativa namnet citonsyracykeln). Från citrat avspjälkas kolatomer i form av CO2 och efter ett varv i Krebscykeln återstår oxaloacetat, som då kan ta hand om en ny molekyl acetyl-CoA.
Bild: Karl-Erik Johansson (BVF, SLU). - Klicka på bilden för att förstora den.
Från en glukosmolekyl, som har 6 kolatomer, bildas under glykolysen två molekyler pyruvat, som har 3 kolatomer vardera. Under aeroba betingelser och om den aktuella bakterien har aerob metabolism, så fermenteras inte pyruvatmolekylen, utan kan istället omvandlas till acetyl-Coenzym A (acteyl-CoA), under det att en molekyl NAD+ reduceras till NADH. Acetylgruppen i acetyl-CoA har två kolatomer eftersom koldioxid (CO2) har spjälkats av och kan nu gå in i Krebscykeln (= citronsyracykeln). Krebscykeln är ett försteg till elektrontransportkedjan (= andningskedjan).
Alla enzymer, som krävs för Krebcykeln finns hos bakterier i cytoplasman (= cytosol), eftersom bakterier saknar mitokondrier. Acetylgruppen binds i första steget i Krebscykeln till oxaloacetat, som har fyra kolatomer och det bildas då citrat, som har sex kolatomer. Efter ett varv i Krebscykeln har 2 molekyler CO2 avspjälkats, 3 molekyler NADH, 1 molekyl FADH2 och 1 molekyl ATP har producerats av varje pyruvatmolekyl. Det som då återstår av citratmolekylen, är återigen oxaloacetat, som kan acetyleras på nytt och snurra ett varv till i cykeln. Totalt har alltså 8 molekyler NADH, 2 molekyler FADH2 och 2 molekyler ATP bildats från 1 molekyl glukos. Egentligen är det inte ATP, som bildas, utan GTP, men GTP omvandlas i cellen till ATP genom en reakton med ADP. NADH och FADH2 utnyttjas i elektrontransportkedjan för syntes av ATP.
Vissa bakterier har ett enzym (tryptofanas), som kan bilda pyruvat från aminosyror (tryptofan). Pyruvat kan sedan metaboliseras i Krebscykeln. Andra bakterier kan bryta ner cystein till substanser som kan gå in i Krebscykeln. För att aminosyror ska kunna brytas ner i Krebscykeln, måste de först deamineras, d.v.s. amingruppen måste omvandlas till ammoniak (NH3) eller snarare till en ammoniumjon (NH4+), som kan utsöndras. Dessutom finns det bakterier, som kan metabolisera lipider till glycerol och fettsyror. Glycerol kan omvandlas till pyruvat och metaboliseras vidare i Krebscykeln och fettsyror kan gå direkt in i Krebscykeln. Det finns alltså många vägar in i Krebscykeln och bakterier med aerob, fakultativt anaerob eller mikroaerofil metabolism kan därför utvinna energi ur många olika typer av substanser.
Uppdaterad: 2019-10-10.
Streptococcer brukar ibland delas in i s.k. Lancefield-grupper efter den amerikanska mikrobiologen Rebecca Lancefield (1895 - 1981), som utveckade ett system för serologisk indelning baserad på kolhydratsammansättningen i cellväggen. De olika grupperna kallas Lancefield group A till V (utom I och J). Dessutom finns en grupp NG (= non-groupable).
Uppdaterad: 2017-09-17.
.")
Schematisk bild av en lipoplysackarid (LPS). Variationer i strukturen förekommer hos olika bakteriearter. Kärnan kan delas upp i en inre och en yttre del. Den inre delen består av 1-3 KDO-rester (keto-deoxyoktulosonat) och 2-3 heptosrester. Den yttre delen består av flera olika hexosrester (t.ex. D-glukos, D-mannos och D-galaktos).
Bild: Karl-Erik Johansson (BVF, SLU) - Klicka på bilden för att förstora den.
Lipopolysackarider (LPS), som också kallas lipoglykaner, finns bara i yttermembranet hos gramnegativa bakterier. LPS är ett s.k. endotoxin (se även toxin nedan) och den toxiska egenskapen sitter i lipid A (se nedan).
LPS är en amfipatisk molekyl, d.v.s. den har båda hydrofila (vattenälskande) och hydrofoba (vattenavstötande = lipidälskande) regioner. Den hydrofoba delen (kolvätekedjor) förankrar LPS i det yttre lipidskikt av bakteriens yttermembran och den hydrofila delen (laddade grupper) pekar utåt mot bakteriens omgivning. Kemiskt består LPS av en lipddel och en polysackariddel. Lipiddelen består av lipid A, som är en fosforylerad glukosamindisackarid med flera (4-6) bundna kolvätekedjor, vilka utgör den hydrofoba delen av molekylen. Kolhydratdelen består av en s.k. kärna ("core oligosackarides", "core antigen" eller R-antigen), som är direkt bunden lipid A. En polysackarid (O-polysackarid, O-antigen eller somatiskt antigen) är bunden till R-antigenet.
Tack vara de olikheter, som finns i LPS från olika gramnegativa bakterier, kan dess antigena egenskaper användas för typning och subtypning av bakterier. De antigena egenskaperna varierar också i olika delar av LPS:
Djur (inkl. människa) utsätts hela tiden för små mängder LPS i blodcirkulationen på grund av omsättningen av gramnegativa tarmbakterier och därför stimuleras hela tiden det medfödda immunsystemet. Om man utsätts för större mängder LPS som t.ex vid en sepsis frisätts cytokiner, vilket leder till feber och inflammation. Vid toxiska koncentrationer av LPS kan blodproppar bildas i kapillärsystemet vilket i sin tur kan leda till livshotande tillstånd.
Uppdaterad: 2020-03-19.
Logaritmer (log) är ett hjälpmedel vid beräkningar av stora tal. När man diskuterar antal mikroorganismer i odlingar från t.ex. livsmedelsprov eller när man gör och beskriver spädningar används oftast logaritmer för att uttrycka spädningsfaktorn.
Logaritmen för ett viss tal (x) är det tal (y), till vilket ett annat tal (b) måste upphöjas, för att ge talet x. D.v.s:
x = by
Talet b kallas för basen i det logaritmsystem man vill använda och vanligast inom natur- och ingenjörsvetenskap är att man använder talet 10 som bas och då talar man om tiologaritmer eller vanliga logaritmer, som brukar skrivas log10 eller lg10. För enkelhets skull skrivs eller sägs ofta bara log eller lg, när man avser tiologaritmen. Ett exempel:
lg 1000 = 3 eftersom 1000 = 10x10x10, som ju också kan skrivas 103.
Enligt logaritmreglerna gäller bl.a. följande för alla typer av logaritmer:
Av andra regeln följer att log10 (1 / b) = 0-log10b = -log10b
D.v.s. logaritmer kan också vara negativa, t.ex:
log10 (1/1000) = -3, eftersom log10 (1/1000) = 0-3 = -3
Alltså:
...
103 = 1000
102 = 100
101 = 10
100 = 1 (kan egentligen inte förklaras matematiskt, men har av praktiska skäl definierats som 1)
10-1 = 0,1
10-2 = 0,01
10-3 = 0,001
...
Inom mikrobiologi vill man ofta göra spädningsserier när man ska bestämma hur många mikroorganismer det finns i ett prov. Vanligast är att man gör en 10-stegs spädningsserie för att uppnå hög noggrannhet vid stora utspädningar. Detta innebär att spädningsfaktorn är 1:10 i varje steg och att man från ett prov tar 1 del provlösning till 9 delar odlingsmedium (eller annan lämplig vätska) i ett provrör, blandar och överför 1 del av blandningen till 9 delar odlingsmedium i ett nytt provrör och så vidare tills man har uppnått lämplig spädningsgrad. Då kommer man att få en serie provrör med spädningsfaktorerna 1:10, 1:100, 1:1000, 1:10 000 o.s.v. Dessa spädningsfaktorer kan också skrivas som: 1/10, 1/100, 1/1000, 1/10 000 o.s.v. eller 0,1, 0,01, 0,001, 0,0001. Detta kan ju också uttryckas med hjälp av logaritmer, d.v.s. 10-1, 10-2, 10-3, 10-4 o.s.v.
En 1:1000 spädning, pratar man därför ofta lite slarvigt om som en minus-tre-spädning.
Läs mer om hur man utnyttjar spädningsserien för att bestämma bakteriemängder under Kolonibildande enhet (CFU).
Logaritmer med andra baser än 10 används av praktiska skäl inom olika discipliner.
Binära logaritmer (lb) med basen 2 används inom bl.a. datorvetenskap, bioinformatik och fotografering.
Naturliga logaritmer (ln) med basen e [ett transcendent tal (gränsvärde) som är ungefär lika med 2,71828 ...] används inom bl.a. matematik, fysik och kemi.
Uppdaterad: 2021-06-23.
.")
Figuren visar blodagarplattor med fyra olika bakterier, som har odlats under 24 timmar vid 37° C. Dessa bakterier (A, Erysipelothrix rhusipathiae; B, Streptococcus agalactiae; C, Staphylococcus aureus; D, Bacillus cereus) representerar fyra olika kolonistorlekar. Längden av skalstrecken motsvarar 5 mm. - Klicka på bilden för att förstora den.
.")
Figuren visar närbilder av bakterierna på ovanstående plattor. Alla fyra bilderna togs från samma avstånd. Längden av skalstrecken motsvarar 5 mm. - Klicka på bilden för att förstora den.
En bakteries koloniutseende på agarplattor är bland annat beroende av tillväxtmedium, tillgång på näring, inkuberingstid och temperatur. Det finns många olika sorters odlingsmedia, som används för typning och/eller selektiv odling av bakterier. Samma bakterieart kan alltså ha olika utseende, beroende på vilket agarmedium den växer på. Vanligen bedöms en bakteries koloniutseende på en blodagarplatta. Enskilda bakteriekolonier bildas oftast från en enda bakterie. När bakteriekolonin blir synlig består den av åtskilliga miljoner bakterieceller. En sådan samling av bakterier brukar kallas för en kolonibildande enhet (CFU), eftersom man inte kan vara alldeles säker på att den bildades av en enda bakteriecell. Bakteriekolonier varierar i utseende vad gäller storlek, färg och form mm. På VetBact klassificeras bakterierna bl.a. efter hur stora kolonier de bildar enligt följande:
Andra makromorfologiska egenskaper hos bakteriekolonien, som kan användas vid identifiering/klassificering, är följande:
Hela bakteriekolonin
Kolonikantens form
Kolonins struktur
Kolonins yta
Kolonins profil
Dessutom kan andra karaktäristika användas för identifiering/klassificering såsom färg, opacitet, lukt och eventuell hemolys.
Uppdaterad: 2018-06-28.
.")
Instrumentet på bilden tillhör Institutionen för Husdjurens biovetenskaper vid Sveriges lantbruksuniversitet (SLU). Marina Falk (018-672389) är ansvarig för utrustningen och man kan få prover analyserade. - Klicka på bilden för att förstora den.
.")
Schematisk beskrivning av principen for MALDI-TOF MS. - Klicka på bilden för att förstora den
Masspektrometri baserad på MALDI-TOF (eller bara Maldi) innebär att bakterie isolatet, som ska analyseras, adsorberas till någon typ av bärarmaterial (matrix). Sedan bestrålas isolatet med laserUVljus, vilket innebär att molekylerna i bakterien slås sönder till positivt laddade fragment (jonisering), som slungas iväg mot en detektor. Tiden det tar för fragmentet att nå detektorn (time of flight) mäts. Tiden är beroende av fragmentets storlek och laddning. Även mycket stora molekyler (proteiner och nukleinsyra) kan fragmenteras och joniseras på detta sätt. Stora molekyler ger upphov till många fragment och ett karakteristiskt masspektrum, som kan användas för identifiering.
Man kan utföra dessa analyser direkt på bakteriekolonier och det erhållna masspektrumet jämförs sedan med lagrade masspektra (referensbibliotek) av kända bakteriearter. Man har alltså ett analyssvar inom någon minut och metoden anses vara mycket tillförlitlig. Ju fler kända bakterier man kan jämföra sina masspektra med, desto säkrare kommer metoden att bli.
MALDI-TOF MS används redan på vissa laboratorier för veterinärmedicinsk bakteriologi och många forskare anser att denna teknik kommer att vara framtidens rutinmetod för identifiering av bakterier. Instrumentet är fortfarande mycket dyrt, men materialkostnader blir låga.
Mjukvaran till Maldi-systemet innehåller en databas med information om vilka fragment man kan tänkas få från en viss bakterie. Denna information kan sedan systemet använda för att jämföra med okända isolat och räkna ut ett poängvärde för varje isolat. Poängvärdet är ett mått på sannolikheten för att ett visst bakterieisolat ska representera en viss bakterieart/stam. Ett poängvärde mellan 0,000 och 1,699 innebär att identifiering inte är möjlig eftersom masspektrat inte matchar till någon bakterie i referensbiblioteket. Ett poängvärde mellan 1,700 och 1,999 betyder att det är troligt att det okända isolatet representerar ett visst släkte men resultatet är inte tillförlitligt på species nivå. Ett poängvärde mellan 2,000 och 3,000 innebär att identifiering av bakterien är med stor sannolikt korrekt på såväl genus som species nivå.
Uppdaterad: 2024-12-09.
.")
Mikroaerob odling med hjälp av kommersiella gasgenererande kuvert i lufttäta odlingsburkar - Klicka på bilden för att förstora den.
Mikroaerofila bakterier (eller mikroaerofiler) kräver en reducerad syrekoncentration för att växa. De har en optimal tillväxt vid 5-10% syre och vissa av dem stimuleras av 1-10% koldioxid. Bakterier som stimuleras av koldioxid kallas för kapnofila bakterier (eller kapnofiler).
Den normala atmosfären består av ca 21% syrgas och denna koncentration är alltså giftig för mikroaerofila bakterier, som troligen har mindre effektiva enzymer (katalas eller peroxidas och superoxiddismutas) för att avgifta väteperoxid och superoxidradikalen, som bildas när syre är löst i vatten (se anaeroba bakterier).
Tillväxten hos vissa arter förbättras dessutom genom närvaro av vätgas, men orsaken till detta är dock inte klarlagd.
Odling av mikroaerofila bakterier kräver särskild odlingsteknik. Flera metoder finns tillgängliga för att uppnå en optimala gasblandning såsom kommersiella gasgenererande kuvert, som öppnas och läggs i lufttäta burkar eller plastpåsar. Man kan också använda inkubatorer eller lufttäta burkar med ventiler för evakuering av den normala atmosfären och återfyllning av en optimal gasblandning.
Campylobacter spp., Borrelia spp. och Helicobacter spp.
Uppdaterad: 2023-04-30.
.")
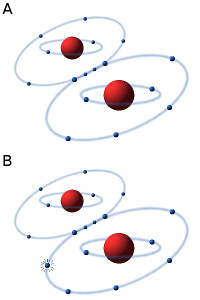
Bilden visar hur strukturen för FAD förändras vid oxidation/reduktion. Notera de två extra väteatomerna i den reducerade formen av FAD. Den kemiska strukturen för AMP och ribos framgår av bilden under ATP i termlistan. - Klicka på bilden för att förstora den.
.")
Bilden visar hur strukturen för NAD förändras vid oxidation/reduktion. Notera den extra väteatomen i den reducerade formen av NAD. Den kemiska strukturen för ADP och ribos framgår av bilden under ATP i termlistan. - Klicka på bilden för att förstora den.
NAD är en dinukleotid, som består av en adenindel och en nikotinamiddel, vilka är sammanfogade genom två fosfatgrupper mellan ribosdelarna på respektive nukleotid. NAD finns i en oxiderad form (NAD+) och en reducerad form (NADH) i alla levande celler och detta är essentiellt för alla levande organismer.
NAD deltar i s.k. redoxprocesser i cellen, d.v.s. kemiska reaktioner där en molekyl reduceras och en annan molekyl oxideras. NAD+ är alltså den oxiderade formen av NAD, som kan oxidera andra molekyler och då själv reduceras till NADH genom att ta upp elektroner (e-). NADH kan i sin tur reducera andra molekyler genom att avge elektroner. Den viktigaste funktionen hos NAD är dessa elektronöverföringsreaktioner. NAD har även andra funktioner, t.ex. som substrat för enzymer, som katalyserar modifiering av makromolekyler ("post translational modification") genom avspjälkning eller bindning av kemiska grupper till t.ex. proteiner.
Den s.k. V-faktorn, som behövs för att vissa bakterier ska växa i artificiella media, utgörs av nikotinamid-adenin-dinucleotide (NAD) eller nikotinamid-adenin-dinukleotidfosfat (NADP, se nedan).
NADP har samma kemiska struktur som NAD samt en extra fosfatgrupp i 2'-ställning på ribosdelen, som hör till adenosin. NADP används av cellen vid bl.a. anabola (biosyntetiska) processer.
FAD består av två huvudkomponenter AMP och flavinmononukleotid (FMN), som kopplade till varandra genom en s.k. fosfodiesterbindning. Nukleotiderna i nukleinsyror (DNA och RNA) är också kopplade till varandra genom sådana bindningar. FMN är ett derivat av riboflavin (= vitamin B2). Tack vare flavingruppen kan FAD fungera som kofaktor till enzymer som katalyserar vissa komplicerade redox-processer, t.ex. i Krebscykeln.
Uppdaterad: 2019-10-31.
Nomenklatur för bakterier avser namngivning och bakterier namnges liksom andra organismer enligt det binomiala system, som infördes av Carl von Linné (1674–1748). Detta innebär att bakterien har ett artnamn, som talar om vilket släkte (genus) den tillhör (genusnamn) och ett namn, som tillsammans med genusnamnet är unikt för bakterien (artepitet). Ett exempel på detta är Moraxella bovis, där genusnamnet talar om att bakterien tillhör genus Moraxella och artartepitetet talar om att bakterien har isolerats från nötkreatur. Genusnamn och artepitet bildar tillsammans det vetenskapliga artnamnet, som alltid skrivs i kursiv stil. Bakterienamn är internationella och man använder latin eller latiniserad grekiska för att bilda namnet. Om missförstånd ej kan uppstå, så kan man förkorta genusnamnet efter att det har skrivits ut första gången i en text, t.ex. M. bovis. Observera dock att det också finns bakterier, som heter Mycoplasmopsis bovis och Mycobacterium bovis.
Det finns strikta internationella regler för hur bakterier ska namnges och dessa regler finns publicerade i en bok som heter: "International Code of Nomenclature of Prokaryotes". För att man ska få ett föreslaget namn accepterat måste man publicera en vetenskaplig artikel om den föreslagna arten och denna måste godkännas av en internationell taxonomikommitté.
Trivialnamn används ofta som ett förenklat sätt att namnge bakteriesläkten. Trivialnamn ska skrivas med liten bokstav och får ej kursiveras. Exempel på trivialnamn är: laktobaciller, mykobakterier, salmonella, stafylokocker och streptokocker. De vetenskapliga namnen för dessa grupper är: genus Lactobacillus (eller Lactobacillus spp.), genus Mycobacterium (eller Mycobacterium spp.), genus Salmonella (eller Salmonella spp.), genus Staphylococcus (eller Staphylococcus spp.) resp. genus Streptococcus (eller Streptococcus spp.).
Om man avser en specifik bakterieart får man inte använda ett trivialnamn som avser ett helt bakteriesläkte.
Ibland finns ett behov av att dela upp bakteriearter i underarter, eftersom de är alltför nära besläktade för att betraktas som olika arter, men alltför avlägset besläktade för att betraktas som samma art. I detta fall införs en underart genom att lägga till ett underartepitet och skriva (subsp. eller ssp.) framför underartepitetet. Ett exempel på detta är Streptococcus equi subsp. equi. När man delar en art i flera underarter, får den ursprungliga arten alltid samma underartepitet som artepitetet.
Ofta vill man också dela in arter och underarter i olika biovarer (biologiska varianter) eller olika stammar, men detta regleras inte lika strikt utan forskare kan själva namnge sina stammar eller biovarer. En typ av biovar är serovar (serologisk variant) där man identifierar olika ytantigener med hjälp av specifika antikroppar. Smittspårning och epidemiologi bygger på att man kan identifiera olika varianter av en och samma bakterieart.
Serovar och serotyp är synonyma och alltså utbytbara termer, men enligt regler i "Bacteriological Code (1990 Revision)", är serovar den term som föredras. Serogrupp är en grupp av bakterier, som innehåller ett gemensamt antigen. En serogrupp kan innehålla flera olika serotyper. Serogrupp är inte en officiell beteckning, men har använts för att klassificera bakterier tillhörande släktena Leptospira, Salmonella, Shigella och Streptococcus.
En bakterieunderart som förekommer i flera tusen olika serovarer är Salmonella enterica subsp. enterica. En vanlig serovar är Dublin och ska man då skriva det fullständiga och korrekta namnet för den bakterien, så blir det Salmonella enterica subsp. enterica serovar Dublin. Observera att namnet på serovaren skrivs med stor bokstav och skall ej kursiveras. Om namnet förekommer på flera ställen i texten kan man skriva S. enterica subsp. enterica serovar Dublin. Även detta är relativt långt att skriva ut och därför har man bestämt att det också är acceptabelt att skriva enbart Salmonella Dublin, utom på första stället i texten där hela namnet måste skrivas ut.
Man kan läsa mer om namngivning av salmonella på VetBact under Salmonella spp. och Salmonella enterica.
Uppdaterad: 2023-09-27.
Med analysparameter inom livsmedelsmikrobiologi avser man olika standardiserade rutinmetoder för bestämning av t.ex. antal långsamväxande bakterier, intestinala enterokocker, odlingsbara mikroorganismer o.s.v.
Uppdaterad: 2013-02-26.
Uppdaterad: 2015-03-03.
.")
Peptidoglykanen i bakteriernas cellvägg är uppbyggd av repeterande enheter bestånde av två aminosackarider, N-acetylglukosamin (NAG) och N-acetylmuraminsyra (NAM). NAM har en oligopeptid, bestående av 4-5 aminosyror (aa), kopplade till sig. Oligopeptiden är också bunden till en annan oligopeptid via en amid-bindning och på så sätt blir alla peptidoglykankedjor sammankopplade till en enda stor jättemolekyl.
Bild: Karl-Erik Johansson (BVF, SLU). - Klicka på bilden för att förstora den.
De flesta grampositiva och gramnegativa bakterier har en peptidoglykan i cellhöljet, som är 20-80 respektive 7-8 nm tjockt. Peptidoglykanen kallas också murein och är en huvudkomponet i de flesta bakteriers cellvägg. Peptidoglykanen är en polymer, som utgör ett glest nätverk i form av en enda stor tredimensionell jättemolekyl, vilken stabiliserar bakteriernas cellhölje.
Peptidoglykanen är uppbyggd av två alternerande kolhydratmolekyler innehållande aminogrupper: N-acetylglukosamin (NAG) och N-acetylmuraminsyra (NAM). Varje NAM-molekyl har har oligopeptid, som består av 4-5 aminosyror, kopplad till sig. Oligopeptiden från den ena peptidogykankedjan är tvärbunden med en annan oligopeptid från en annan peptidoglykankedja (se figuren) via en s.k. amidgrupp (-CO-NH-). Aminosyror (utom glycin) kan existera i två olika isomera former, L-form och D-form, som är spegelbilder av varandra. Levande organismer innehåller vanligtvis bara L-formen av aminosyror, men oligopeptiden i bakteriers cellväggar består av både L- och D-form av de ingående aminosyrorna samt av en aminosyra (meso-diaminopimelinsyra), som inte ingår i vanliga proteiner. Olika arter av bakterier kan ha lite olika sammansättning av oligopeptiden.
Förutom att vara ett mekaniskt skydd och ett skydd mot osmotisk lysering har cellväggen en viktig funktion vid celldelningen. Bakterien sträcker ut sig på längden före celldelningen och då måste cellväggen växa genom nysyntes av peptidoglykanen. När bakteriecellen sedan delar sig, måste peptidoglykanen omformas så att de båda dottercellerna försluts.
Uppdaterad: 2020-04-02.
Uppdaterad: 2013-03-06.
Pyogena bakterier kan orsaka pyemi (se sepsis nedan).
Uppdaterad: 2012-12-12.
Inledning
"Quorum sensing" är ett system för signalering och svar i en population av organismer (t.ex. bakterier), som regleras av populationens storlek. Man kan säga att genuttrycket (= proteinsyntesen) av vissa proteiner kan regleras som svar på en förändring av populationstätheten. Kvorum kommer från latinets quorum "av dem" och syftar på det minsta antal närvarande, som kräves för att en församling ska vara beslutsmässig.
Varför kommunicerar bakterier med varandra?
Om väldigt få bakterier finns närvarande inom ett visst område är det slöseri med energi att tillverka och utsöndra t.ex. vissa enzymer och därför regleras genuttrycket genom quorum sensing. Även bildning av biofilm regleras av quorum sensing. Syntes av beståndsdelarna i en biofilm initieras när den bakteriella populationstätheten har nått en viss nivå.
Hur kommunicerar bakterier med varandra?
Bakterier använder quorum sensing för att vid behov reglera genuttrycket i förhållande till populationens storlek. Detta uppnås genom att bakterien skickar ut kemiska signalsubstanser i form av komplicerade organiska molekyler (polypeptider i grampositiva och N-acyl homoserin laktoner i gramnegativa bakterier). I en population av planktoniska bakterier (se biofilm) blir inte koncentrationen av signalsubstanser tillräckligt hög för att bakterierna ska kunna kommunicera med varandra, men i en biofilm är den det och därför kan de kommunicera.
Länkar
Uppdaterad: 2021-01-21.
.")
Renodling av en bakterie i blandflora med minst fyra olika bakterie species - Klicka på bilden för att förstora den.
Ett kliniskt prov innehåller sällan bakterier i renkultur, utan de föreligger vanligtvis i blandflora. Vid en bakteriell infektionssjukdom hittar man oftast den patogena bakterien i blandning med normalfloran. För att kunna identifiera viktiga bakterier i kliniska prover måste de renodlas (se figur), d.v.s man måste ha dem i renkultur. En renkultur innehåller alltså bara en enda bakterieart och detta kan åstadkommas genom renstrykning (se nedan) på agarplattor.
Uppdaterad: 2025-05-13.
 Renstrykning är en teknik, som används inom bakteriologi för att isolera enskilda bakteriearter eller stammar från ett prov, som innehåller en blandflora av bakterier. Man använder en steril plastögla (eller platinaögla), som har doppats i provet, och stryker ut på 1/3 (alt. 1/4) av en agarplatta. Sedan tar man en ny ögla och stryker från första tredjedelen in i nästa tredjedel av plattan. Detta upprepas sedan en gång (alt. två gånger) till. På detta sätt får man en signifikant minskning av antalet bakterier mellan varje tredjedel (alt. fjärdedel) och chansen är god att man någonstans på plattan kan hitta enskilda kolonier av den eller de bakterier man vill isolera. Bilden till höger visar en renspridning av Staphylococcus aureus subsp. aureus i tre steg på en nötblodagarplatta. De båda bilderna (A och B) visar samma platta fotograferad med belysning ovanifrån respektive i motljus. Notera att på platta B kan man se spåren av stryken även om inga bakteriekolonier kan observeras.
Renstrykning är en teknik, som används inom bakteriologi för att isolera enskilda bakteriearter eller stammar från ett prov, som innehåller en blandflora av bakterier. Man använder en steril plastögla (eller platinaögla), som har doppats i provet, och stryker ut på 1/3 (alt. 1/4) av en agarplatta. Sedan tar man en ny ögla och stryker från första tredjedelen in i nästa tredjedel av plattan. Detta upprepas sedan en gång (alt. två gånger) till. På detta sätt får man en signifikant minskning av antalet bakterier mellan varje tredjedel (alt. fjärdedel) och chansen är god att man någonstans på plattan kan hitta enskilda kolonier av den eller de bakterier man vill isolera. Bilden till höger visar en renspridning av Staphylococcus aureus subsp. aureus i tre steg på en nötblodagarplatta. De båda bilderna (A och B) visar samma platta fotograferad med belysning ovanifrån respektive i motljus. Notera att på platta B kan man se spåren av stryken även om inga bakteriekolonier kan observeras.
En koloni, som kommer från en renstrykning, utgör i allmänhet en klon eftersom den med stor sannolikhet härstammar från en enda bakteriecell. Det finns dock undantag eftersom vissa bakterier växer i form av aggregat snarare än enskilda celler. Om man arbetar med bakterier, som har en tendens att svärma, får man försöka använda plattor, som minskar den risken (t.ex. CLED-agar för vissa bakterier).
Uppdaterad: 2013-03-05.
Många bakterier är rörliga och de kan förflytta sig genom att utnyttja principiellt skilda mekanismer. Rörelsemekanismer har utvecklats för att bakterier ska kunna attraheras eller repelleras av vissa stimuli. Rörlighet är en egenskap, som används för karaktärisering och identifiering av bakterier och därför har man utvecklat metoder för att påvisa rörlighet. Bakteriers rörelseförmåga måste kunna skiljas från s.k. Brownsk molekylrörelse, som är slumpmässig och beror på att värmerörelse hos vattenmolekyler, som kan knuffa till bakterier så att det uppfattas som om bakterierna rör sig under mikroskopet.
Videoklipp som visar rörligheten hos Brachyspira murdochii (Spirochaetota).
Bakterier kan ha flageller, som kan vara en eller flera till antalet, och som fungerar som propellrar och gör att bakterierna kan röra sig i en bestämd riktning. Mycket förenklat kan man säga att flagellen består av ett rörligt filament, som är kopplat till en molekylär motor (basalkropp) i bakteriens hölje via en krok. Flagellen består av många subenheter av proteinet flagellin, som bildar en ihålig och flexibel cylinder. Den molekylära motorn, som är uppbyggd av proteinsubenheter, drivs av protongradienten (eller en Na+-jongradient) över plasmamembranet och kan få filamenten att rotera medsols eller motsols. Denna rotation får bakterierna att simma i en bestämd riktning respektive tumla. Riktningen beror av yttre stimuli och vid tumlandet kan bakterierna byta rörelseriktning.
Bakterier inom fylumet Spirochaetota har s.k. periplasmiska flageller (= endoflageller eller axialfilament), som är lokaliserade till det plasmatiska utrymmet mellan cellmembranet och yttermembranet. Detta arrangemang resulterar i en skruvliknande rörelse (eller flatvågsrörelse), som gör att dessa bakterier kan ta sig fram i mycket viskösa material (t.ex. mucus).
Bakterier, som har så kallade typ IV pili kan röra sig genom att de yttre ändarna av dessa pili, som har hakar, vidhäftar till ett fast substrat, som kan utgöras av en yta som bakterien sitter på eller andra bakterier. När piluset kontraheras, dras bakterien framåt. Rörelse, som produceras av typ IV-pili är typiskt ryckig och den kallas helt enkel för ryckig rörelse. Pseudomonas aeruginosa uppvisar ryckig rörelse.
Vissa bakterier kan glida på våta ytor, men de molekylära mekanismerna för detta är ofullständigt utredda. Exempel på bakterier, som har denna förmåga är: medlemmar av fylumet Cyanobacteria samt släktena Flavobacteria och Mycoplasma.
Vissa patogena bakterier kan röra sig inuti värdcellen genom att utnyttja dess cytoskelett. Cytoskelettet används normalt för att flytta organeller inuti eukaryota cellen. Genom att stimulera aktinpolymerisering vid en av sina poler, kan dessa bakterier bilda en slags svans, som driver dem genom värdcellens cytoplasma. Exempel på bakterier, som kan använda denna mekanism, är arter inom släktena Burkholderia, Listeria, Rickettsia och Shigella.
Rörlighetstester kan göras i provrör innehållande ett "halvfast" (semisolid) medium, d.v.s. ett medium med låg koncentration av agar. Man ympar mediet med en plastögla, som sticks ner i agarn och dras upp igen. Om bakterien är rörlig så grumlas allt medium i röret, men om den är orörlig får man bara grumling i själva sticket. Bilden visar en negativ kontoll (A), Staphylococcus aureus subsp. aureus (B) som är orörlig samt Escherichia coli (C), som är rörlig.
Uppdaterad: 2025-07-01.
Salmonella spp. har, liksom andra medlemmar av familjen Enterobacterioceae, tre huvudtyper av antigener, som kan användas vid serologisk diagnostik och identifiering. Dessa antigener är speciellt användbara för identifiering av bakterier inom följande taxa: Escherichia coli, Klebsiella spp., Salmonella spp. och Shigella spp. För epidemiologiska studier delar man in Salmonella enterica subsp. enterica i olika serovarer (= serovarianter) beroende på vilken uppsättning av antigener stammen ifråga bär på. Ordet serotyp används också synonymt med serovar. Man har definierat följande tre huvudtyper av antigener:
Kauffman-Whites klassificeringsschema är baserat på O- och H-antigener hos Salmonella spp. Man har identifierat fler än 2500 serovarer (serotyper) baserade på O- och H-antigener hos Salmonella spp. För epidemiologiska studier testas Salmonella-isolat först med ett polyvalent O-antiserum riktat mot alla olika serotyper. Om isolatet är positivt, så testas det sedan med monovalenta O-antisera riktade mot specifica serotyper. Om isolatet är negativt med polyvalent antiserum, så måste det värmebehandlas för att förstöra ev. K-antigen, som kan maskera O-antigener. Därefter upprepas testet med det polyvalenta antiserumet och om det då är positivt, så måste man också gå vidare med monovalenta antisera. På motsvarande sätt testar man också vilka typer av H-antigener, som stammen i fråga bär på. O-antigenet bestämmer vilken serogrupp ett visst Salmonella-isolat representerar och i kombination med H-antigenet kan serovaret (= serotypen) definieras.
Exempel: En salmonella-typ som heter Salmonella enterica subsp. enterica Serovar Dublin har antigenuppsättningen: 1,9,12:g,p:inga fas 2, vilket betyder att den har:
De antikroppar, som används i testet är kopplade till mycket små latex-kulor, för att latexkulorna ska bilda synliga aggregat med bakterierna och det är detta som kallas för agglutination. En bakterie, som är positiv i agglutination med polyvalenta antisera mot både O- och H-antigenet representerar med stor sannolikhet ett Salmonella-isolat, men för att kunna göra epidemiologiska studier (smittspårning) måste man bestämma vilket serovar isolatet tillhör.
Hänvisning: Läs mer om salmonellornas nomenklatur på bakteriesidorna om Salmonella spp. och Salmonella enterica på VetBact och om olika serovarer i Termlistan på VetBact.
Uppdaterad: 2023-02-22.
Släktet Salmonella uppdelas i två (eller tre) arter: Salmonella bongori, Salmonella enterica (och ev. Salmonella subterranea), se även Salmonella spp. Salmonella enterica indelas i sex underarter, där framför allt Salmonella enterica subsp. enterica är av intresse inom human- och veterinärmedicin. S. enterica subsp. enterica indelas i sin tur i flera hundra olika serovarer, baserade på deras antigensammansättning. Förkortningen S. för Salmonella kan man använda om det i texten inte förekommer några tvetydigheter. De olika serovarerna får då väldigt långa namn, så det finns då en konvention att man förkortar dem på ett visst sätt, där Salmonella bör skrivas ut och i kursiverad stil. Namnet på serovaret ska skrivas i normal stil och med stor första bokstav. Första gången det långa vetenskapliga namnet används i en text, ska det dock skrivas ut. I tabellen nedan listas några vanligt förekommande serovarer med fullständigt och förkortat namn samt deras vanligaste värddjur.
Tabell:
| Vetenskapligt namn | Förkortat namn* | Djurslag |
| S. enterica subsp. enterica Serovar Anatum | Salmonella Anatum | Nötkreatur |
| S. enterica subsp. enterica Serovar Cholerasuis | Salmonella Cholerasuis | Gris, vildsvin |
| S. enterica subsp. enterica Serovar Derby | Salmonella Derby | Gris |
| S. enterica subsp. enterica Serovar Diarizonae | Salmonella Diarizonae | Får |
| S. enterica subsp. enterica Serovar Dublin | Salmonella Dublin | Nötkreatur |
| S. enterica subsp. enterica Serovar Enteritidis | Salmonella Enteritidis | Fjäderfä, igelkott, människa |
| S. enterica subsp. enterica Serovar Gallinarum | Salmonella Gallinarum | Fjäderfä |
| S. enterica subsp. enterica Serovar Hadar | Salmonella Hadar | Fjäderfä |
| S. enterica subsp. enterica Serovar Kentucky | Salmonella Kentucky | Fjäderfä |
| S. enterica subsp. enterica Serovar Livingstone | Salmonella Livingstone | Fjäderfä |
| S. enterica subsp. enterica Serovar Newport | Salmonella Newport | Fjäderfä, gris, nötkreatur, vattenfåglar |
| S. enterica subsp. enterica Serovar Paratyphi | Salmonella Paratyphi | Människa |
| S. enterica subsp. enterica Serovar Pullorum | Salmonella Pullorum | Fjäderfä |
| S. enterica subsp. enterica Serovar Senftenberg | Salmonella Senftenberg | Kalkon |
| S. enterica subsp. enterica Serovar Typhi | Salmonella Typhi | Människa |
| S. enterica subsp. enterica Serovar Typhimurium | Salmonella Typhimurium | Gris, fjäderfä, katt, nötkreatur, igelkott, människa |
------------------------------------------------------------------------
*Man kan även förkorta Salmonella till S. här, om det inte kan missförstås, även om det inte ser så snyggt ut i text.
Observera att det vanligaste serovaret på ett visst djurslag och i ett visst geografiskt område, inte nödvändigtvis är det vanligaste serovaret på samma djurlag, men i ett annat geografiskt område.
Uppdaterad: 2023-04-05.
I bakterier har det skett en fantastisk evolution av sekretionssystem för att transportera substanser över cellhöljet (= plasmamembran, cellvägg och ev. yttermembran). Bakteriella sekretionssystem är "nanomaskiner" i form av proteinkomplex lokaliserade till cellmembranen. Hos patogena bakterier är deras funktion att utsöndra olika substanser, s.k. effektormolekyler, som huvudsakligen är proteiner, vilka gör att bakterien sedan kan kolonisera eller invadera värdcellen. Effektormolekylerna kan också manipulera värdcellen så att den får sämre förmåga att överleva bakterieinfektionen, därför att de utgör toxiner, adhesiner, eller nedbrytningsenzymer mm. Sekretionssystemen är alltså mycket viktiga virulensfaktorer och de kan indelas i olika typer beroende på deras sammansättning, struktur, mekanism och evolutionärt släktskap. Gramnegativa och grampositiva bakterier kan ha (minst) åtta respektive (minst) fyra olika sekretionssystem och det finns två principiellt olika mekanismer (enstegs- och tvåstegs-mekanismen).
Genom enstegsmekanismen transporteras effektormolekylerna direkt över bakteriens cellmembran(er) in i värdcellen och det kan ske i båda gramnegativa och grampositiva bakterier. Genom tvåstegsmekanismen transporteras effektormolekylerna först genom plasmamembranet till bakteriens periplasma och sedan med hjälp av andra proteinkomplex ut genom yttermembranet och in i värdcellen. Detta kan bara ske i gramnegativa bakterier.
Sekretion över plasmamembranet sker i både grampositiva och gramnegativa bakterier och det finns tre huvudsystem för denna typ av transport:
Sekretion över yttermembranet behöver givetvis bara ske i gramnegativa bakterier och genom antingen enstegsmekanismen eller tvåstegsmekanismen.
Man brukar dela in sekretionssystemen i följande sju olika huvudtyper, men denna indelning är inte fullständig och ibland inte heller helt klar. Sedan finns det också undertyper av dessa huvudtyper:
Man kan hitta fina bilder på de olika sekretionssystemen på webbplatsen SlideShare (www.slideshare.net) om man söker på "bacterial secretion system".
Uppdaterad: 2020-02-27.
Termen septikemi används ibland som en synonym till sepsis och avser då en sepsis där man har påvisat bakterieväxt i blodet.
Enbart förekomst av bakterier i blodet är inte synonymt med sepsis utan detta tillstånd kallas bakteriemi och är övergående.
Pyemi är en form av sepsis, som resulterar i omfattande bölder av metastatisk natur. Pyemi orsakas oftast av närvaro av pyogena (varbildande) bakterier i blodet.
Uppdaterad: 2012-12-11.
Ordet siderofor kommer från grekiskan och betyder järnbärare. Sideroforer är lågmolekylära, substanser, som har mycket hög affinitet för järn (Fe3+) och de binder järn genom s.k. kelatbindning. Järn är essentiellt för nästan alla livsformer, därför att det är en nödvändig komponent i andningskedjan och vid DNA-syntes. Många bakterier kan utsöndra sideroforer, som gör att de kan ta upp järn även i en omgivning där koncentration av fritt järn är mycket låg. Sedan kan järnet tas upp av bakterien eftersom den har receptorer för sideroforer på cellytan. Hos däggdjur är järn hårt bundet till olika proteiner (t.ex. ferritin, laktoferrin, hemoglobin och transferrin) och därför behöver patogena bakterier binda upp järn i form av lösliga komplex med sideroforer. Sideroforer betraktas som virulensfaktorer och år 2015 hade ca 300 olika sideroforer från olika bakterier beskrivits.
Exempel på sideroforer
| Bakterie | Siderofor |
| Bacillus anthracis | Bacillibaktin |
| Bacillus subtilis | Bacillibaktin |
| Burkholderia pseudomallei | Malleobaktin |
| Escherichia coli och några andra enterobakterier ... | Enterobaktin |
| ... t.ex. Salmonella spp. | Enterobaktin |
| Några arter inom släktet Mycobacterium (t.ex. M. tuberculosis) | Mykobaktin |
| Pseudomonas aeruginosa | Pyokelin och pyoverdin |
| Yersinia enterocolitica | Yersiniabaktin |
| Y. pestis | Yersiniabaktin |
| Y. pseudotuberculosis | Yersiniabaktin |
Se även individuella bakteriesidor för fler exempel.
Uppdaterad: 2017-11-10.
Inom både veterinär- och humanmedicin brukar man karaktärisera infektionssjukdomar (orsakade av bakterier, parasiter, svampar eller virus mm) efter olika kriterier och här följer några exempel på indelningar av infektionssjukdomar inom dessa båda discipliner.
Enzooti motsvarar endemi inom humanmedicin och en enzootisk sjukdom är en sjukdom, som alltid finns i en viss djurpopulation, men som vid en viss tidpunkt bara påverkar ett mindre antal djur. Enzootisk pneumoni hos gris, som orsakas av Mycoplasma hyopneuminiae, är ett exempel på en bakterieorsakad enzooti.
Epizooti motsvarar epidemi inom humanmedicin och en epizootisk sjukdom, är en allvarlig djursjukdom med utbredd smittspridning. Mjältbrand, som orsakas av Bacillus anthracis, är exempel på en bakterieorsakad epizooti.
Panzooti syftar på en allvarlig infektionssjukdom, som sprids över stora delar av världen (en eller flera kontinemter) och drabbar ett eller flera djurslag. Afrikansk svinpest är ett exempel på en virusorsakad panzooti. Inom humanmedicinen talar man istället om pandemi. Kolera, som orsakas av bakterien Vibrio cholerae gav upphov till tre pandemier på 1800-talet.
Zoonos är ett begrepp, som används inom både veterinär- och humanmedicin, eftersom det syftar på infektionssjukdomar, som kan överföras mellan djur och människor via livsmedel, direktkontakt med smittbärande djur eller genom indirekt kontakt med smittbärande djur via t.ex. insektsbett. Brucellos, campylobakterios, nötkreaturstuberkulos och salmonellos är exempel på zoonoser, som orsakas av följande zoonotiska bakterier: Brucella melitensis, Campylobacter jejuni, Mycobacterium bovis resp. Salmonella enterica subsp. enterica.
Uppdaterad: 2024-02-08.
Sporer hos bakterier är inte samma sak som sporer hos svampar. Bakterier använder inte sporer för att föröka sig utan för att överleva ogynnsamma betingelser (brist på näringsämnen, extremt pH, hög temperatur mm). Vissa bakterier kan alltså övergå till ett vilostadium genom att bilda sporer.
Endosporer bildas, som namnet antyder, inuti baktericellen. Endosporer är mycket motståndskraftiga och kan bildas av medlemmar av fylumet Bacillota. De släkten, som har störst betydelse inom veterinärmedicinen är Bacillus och det närbesäktade släktet Paenibacillus samt Clostridium och de närbesläktade släktena Paeniclostridium och Clostridioides. I varje bakteriecell bildas endast en spor och den kan överleva utan tillgång på näringsämnen. Endosporer tål höga och låga temperaturer, intorkning, kemiska desinfektionsmedel och UV-strålning och anledningen till att de är så resistenta är att de innehåller nästan inget vatten och att cellväggen innehåller dipikolinsyra. Sporen innehåller också mycket calcium (Ca) och när Ca2+ pumpas in i sporen, så pumpas vatten ut. Förutom höljet (cortex) innehåller sporen i stort sett bara DNA, ribosomer och polymeraser. Detta är tillräckligt för att sporerna ska kunna germinera, d.v.s. utvecklas till bakterier igen, när förhållandena har blivit gynnsamma.
Eftersom endosporer är så resistenta, krävs det behandling med fuktig värme (+121°C) under 15 min för att dessa sporer skall avdödas.
Endosporer har betydelse för identifiering av bakterier. Läget av sporen i modercellen kan ge information om tänkbara bakterier. B. cereus, B. subtilis och C. tetani t.ex. har central, subterminal respective terminal spor.
Videoklipp på YouTube som rekommenderas: "Bacterial Spore Formation".
Ordningarna Nostocales och Stigonematales inom fylumet Cyanobacteria bildar så kallade akineter, som är en typ av vilostadium för dessa bakterier. Akineter har inte så stor likhet med endosporer och de är inte alls lika resistenta, men i form av akineter kan dessa cyanobakterier överleva kyla (vintertid) och intorkning.
Släktena Actinomyces och Streptomyces, som hör till fylumet Actinomycetota kan bilda en typ av sporer, som ibland kallas exosporer. Exosporer uppkommer genom avknoppning av de mycelliknande strukturer (filament), som bildas när dessa bakterier växer. Exosporer är inte alls lika resistenta som endosporer och har därför inte samma kliniska betydelse. De har också en helt annan uppbyggnad.
Är egentligen något annat än sporer, se VBNC (Viable but nonculturable) nedan.
Uppdaterad: 2023-03-08.
.")
Proteus vulgaris, på blodagarplatta inkuberad i 24 h, vid 37 °C. Kolonierna svärmar och sprider ut sig till en matta på agarplattan.
Bild: Ingrid Hansson (BVF, SLU) & Lise-Lotte Fernström (BVF, SLU).
En bakterie, som svärmar, bildar inte diskreta kolonier på en agarplatta, utan flyter ut över hela agarytan med en hastighet på 2–10 μm/s. Om en bakterie svärmar, kan man inte bestämma dess makromorfologi och inte heller räkna kolonier. Man kan dock förhindra svärmning av vissa bakterier genom att odla dem på ett medium med låg elektolythalt (= låg saltkoncentration), som t.ex. CLED agar. En variant av MacConkey-agar utan NaCl kan också användas för att undvika svärmning. Bakterier, som kan svärma hittar man ofta inom släktena: Aeromonas, Bacillus, Brachyspira, Campylobacter, Clostridium, Escherichia, Paeniclostridium, Proteus, Pseudomonas, Salmonella, Serratia, Vibrio och Yersinia. Man vet inte varför vissa bakterier svärmar, men det har föreslagits att det är för att snabbt förflytta sig till områden med mindre konkurrens om näringsämnen. Svärmande bakterier har vanligen fler flageller per ytenhet än vad planktoniska (fritt svävande) bakterier har. Det har observerats att aktivt svärmande bakterier av Salmonella Typhimurium uppvisar en förhöjd resistens mot vissa antibiotika i jämförelse med icke svärmande celler.
Uppdaterad: 2020-01-27.
| Rang | Ändelse | Exempel |
| Underart | - | Campylobacter jejuni subsp. jejuni |
| Art | - | Campylobacter jejuni |
| Släkte (Genus) | - | Campylobacter |
| Familj | -aceae | Campylobacteraceae |
| Ordning | -ales | Campylobacterales |
| Klass | -ia | Campylobacteria |
| Fylum | -ota | Campylobacterota |
| Domän | - | Bacteria |
| Alla levande organismer | - | - |
Med bakteriers taxonomi menar man namngivning och klassificering i olika hierarkiska grupper med ökad omfattning. D.v.s. ett släkte kan bestå av flera olika arter, en familj kan bestå av flera olika släkten o.s.v. Man kan oftast känna igen rangen på dess ändelse, d.v.s. slutar ordet på t.ex. -ales, vet man att det är ordningen, som avses. Detta har dock inte blivit konsekvent infört för alla bakteriearter. Med bakteriers fylogeni menar man deras naturliga släktskap, d.v.s. deras evolutionshistoria.
Alla levande organismer brukar delas in i tre domäner: Eukaryoter (Eukarya), bakterier (Bacteria) och arkéer (Archaea). Det finns även andra sätt att dela in organismer, som t.ex. i sju olika riken: Bakterier (Bacteria), arkéer (Archaea), protozoer (Protozoa), kromister (Chromista), växter (Plantae), svampar (Fungi) och djur (Animalia). Vissa bakteriegrupper delas också in i underfamiljer, underordningar och/eller underklasser. Idag har de flesta forskarna (åtminstone bakteriologer) accepterat indelningen av organismer i tre olika domäner och det var Carl Woese (den molekylära fylogenins fader), som införde detta system år 1990 (se ref. 1 nedan). Nu strävar man efter att bakeriernas taxonomi ska återspegla dears evolutionära historia (fylogeni) och allt eftersom man får ökad kunskap om bakterier och deras fylogeni kommer deras taxonomi att revideras.
.")
Med bakteriers fylogeni menar man alltså deras naturliga släktskap, d.v.s. deras evolutionära historia. När man började dela in bakterier i olika taxonomiska kategorier, så var indelningen baserad på ett fåtal egenskaper (t.ex. gramfärgning, biokemiska tester, sporbildningsförmåga, patogenicitet m.m. m.m.). Ju fler egenskaper, som utnyttjade, desto bättre återspeglade indelningen bakteriernas fylogeni. Dessa egenskaper utgjorde dock endast trubbiga verktyg för att studera bakteriers fylogeni. Det verkliga genombrottet kom när man började utnyttja DNA-sekvensdata för att studera bakteriers släktskap. Det var länge standard att basera fylogenetiska träd på sekvensdata från 16S rRNA-gener. Idag har det dock blivit så billigt att göra helgenomsekvenseringar av bakterier, så att man istället basera sina fylogenetiska träd på sekvensdata från stora delar av genomet. Då har man tillgång till upp till 10 000 gånger mer data jämfört med sekvensdata från enbart 16S rRNA-genen, eftersom varje nukleotidposition kan betraktas som en unik och väldefinierad karaktär.
Fylogenetiska träd (eller släktskapsträd), baserade på sekvensdata, beräknas med hjälp av speciella datorprogram, som kan hantera stora mängder data. I den typ av fylogenetiska träd, som visas på VetBact, är de horisontella avstånden proportionella mot sekvenskillnaderna i de jämförda generna. Alltså blir avstånden mellan två organismer kortare, ju närmare de är släkt med varandra.
Reglerna för namngivning av bakterier [International Code of Nomenclature of Prokaryotes (ICNP)] har tidigare inte omfattat den taxonomiska kategorin (rangen) fylum, men nyligen (februari 2021) har medlemmarna av International Committee on Systematics of Prokaryotes (ICSP) beslutat att så ska ske i fortsättningen. Man har bestämt att alla namn på fyla ska skrivas med kursiv stil (vilket har gjorts på VetBact även tidigare) och ha ändelse -ota. Dessutom ska fylumnamnet vara baserat på ett genus, som ska utgöra nomenklaturtypen (se ref. 2 nedan).
Man brukar säga att bakteriernas fyla utgör deras huvudutvecklingslinjer och nu finns det över 40 beskrivna fyla. Elva av dessa fyla, finns representerade i VetBact. Det största fylumet är Pseudomonadota (tidigare Proteobacteria) och det är också det fylum, som innehåller flest patogena bakterier. I VetBact finns för närvarande 92 bakteriearter, som tillhör fylumet Pseudomonadota.
1. Woese CR, Kandler O & Wheelis ML. (1990). Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. PNAS 87(12):4576–4579.
2. Oren A, & Garrity GM (2021). Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 71(10):005056.
Uppdaterad: 2023-04-17.
.")
Fig. 136:1. Mollikuternas (Mykoplasmernas) nya taxonomi. Röda taxa representerar nya namn. Gröna namn representerar taxa där namnen inte har ändrats och som tillhör samma högre taxon som tidigare. Gula namn representerar taxa där namnen inte har ändrats, men som har placerat inom ett annat högre taxon än tidigare. Veterinärmedicinskt viktiga släkten är understrukna. Observera att alla högre taxa inom fylumet Mycoplasmatota ej finns med i denna figur. - Klicka på bilden för att förstora den.
.")
Fig. 136:2. Arter av mollikuter (mykoplasmer) med betydelse inom veterinärmedicin. Överst i varje ruta står namnet på de olika släktena och under släktnamnet följer de olika artnamnen. Samma färgkodning används i Fig. 136:2 som i Fig. 136:1. Alla arter i figuren finns med på VetBact, men A. laidlawii har dock inte så stor veterinärmedicinsk betydelse. - Klicka på bilden för att förstora den.
Mollikuternas (mykoplasmernas) taxonomi har tidigare varit kontroversiell eftersom den inte har återspeglat deras fylogeni d.v.s. deras naturliga släktskap. Nu har en kanadensisk och en israelisk forskargrupp gjort en omfattande studie baserad på helgenomsekvenseringar där man också har föreslagit en genomgripande revidering av mollikuternas taxonomi (se referens 1 nedan). Senare har man också infört nya fylumnamn och mollicuterna, hör nu till fylumet Mycoplasmatota, som tidigare hette Tenericutes (se referens 2 nedan).
I den reviderade taxonomin har man infört en ny ordning, nämligen Mycoplasmoidales, som omfattar de nya familjerna Mycoplasmoidaceae och Metamycoplasmataceae. Familjen Mycoplasmoidaceae innehåller de nya släktena Malacoplasma och Mycoplasmoides samt släktena Eperythrozoon och Ureaplasma. Släktet Eperythrozoon försökte man tidigare överföra till släktet Mycoplasma, när man förstod att dessa organismer inte är rickettsier. Genusnamnet Eperythrozoon har dock prioritet och därför bör dessa organismer fortfarande ha kvar det ursprungliga genusnamnet, trots att de tillhör klassen Mollicutes. Familjen Metamycoplasmataceae innehåller de nya släktena Mesomycoplasma, Metamycoplasma och Mycoplasmopsis.
De enda tidigare kallade mykoplasmerna, som fortfarande hör till ordningen Mycoplasmatales, är de som tillhör den s.k. mycoides-gruppen. Detta är bra, eftersom man i den gruppen hittar de viktigaste "mykoplasmerna" ur veterinärmedicinsk och global synvinkel. Nu ska man alltså bara använda begreppet "mykoplasma" som trivialnamn om man avser organismer, som hör till det reviderade genuset Mycoplasma. I annat fall, bör man använda begreppet "mollikuter". Släktena Spiroplasma, Entomoplasma och Mesoplasma hörde tidigare till ordningen Entomoplasmatales. För att betona det nära släktskapet med mycoides-gruppens mykoplasmer, bör de dock placeras under ordningen Mycoplasmatales. Släktena Spiroplasma, Entomoplasma och Mesoplasma har knappast någon betydelse inom veterinärmedicinen, men egentligen borde de två senare kanske placeras inom de två familjer (Entomoplasmataceae resp. Mesoplasmataceae) under ordningen Mycoplasmatales.
Den nya taxonomin för mollikuterna kommer nog att ställa till med en del missförstånd och orsaka förvirring eftersom de fem nya släktnamnen är så lika varandra. På lite längre sikt kommer dock den nya taxonomin att vara ett stöd för de som är intresserade av sambanden mellan de olika organismerna inom denna intressanta bakteriegrupp.
1. Gupta RS, Sawnani S, Adeolu M et al. 2018. Phylogenetic framework for the phylum Tenericutes based on genome sequence data: proposal for the creation of a new order Mycoplasmoidales ord. nov., containing two new families Mycoplasmoidaceae fam. nov. and Metamycoplasmataceae fam. nov. harbouring Eperythrozoon, Ureaplasma and five novel genera. Antonie van Leeuwenhoek 111:1583–1630.
2. Oren A & Garrity GM. 2021. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 71(10):005056.
Uppdaterad: 2023-04-12.
Bakterier förökar sig genom delning (två identiska dotterceller bildas vid celldelningen). Om tillgången på näringsämnen är konstant och om de fysikal-kemiska egenskaperna hos odlingsmediet inte förändras, så delar de sig med samma hastighet hela tiden. Detta innebär att tillväxten är exponentiell. Den exponentiella tillväxtfasen (log-fasen) kan dock inte fortgå hur länge som helst eftersom odlingsmediet utarmas på näringsämnen och mediets pH förändras vanligtvis. När man startar en odling i flytande medium, tar det lite tid innan bakterierna kommer igång att växa, speciellt från kliniska prover eller från nerkylda kulturer och denna fas av odlingen brukar kallas lag-fas.
Den tid det tar för antalet bakterier att fördubblas under odling kallas generationstid och den kan variera väldigt mycket för olika bakterier. Escherichia coli, som odlas under optimala betingelser har en generationstid på 20 min, medan Mycobacterium avium subsp. paratuberculosis har en generationstid på ca 24 tim. Detta innebär i praktiken att om man sprider dessa två bakterier på lämpliga odlingsplattor, så kan man lätt se kolonier av E. coli efter ett dygn, medan kolonier av M. avium subsp. paratuberculosis inte kan observeras förrän tidigast om 3 månader!
 Bakteriell tillväxt kan beskrivas i en s.k. tillväxtkurva med fyra olika faser: lag-fas, log-fas, stationär fas och deklinationsfas (= dödsfas) (markerade A, B, C respektive D i figuren). Notera att skalan på y-axeln är logaritmisk. Under den stationära fasen dör bakterier med ungefär samma hastiget, som de återbildas vid celldelningarna och i deklinationsfasen dör bakterierna snabbare än de återbildas.
Bakteriell tillväxt kan beskrivas i en s.k. tillväxtkurva med fyra olika faser: lag-fas, log-fas, stationär fas och deklinationsfas (= dödsfas) (markerade A, B, C respektive D i figuren). Notera att skalan på y-axeln är logaritmisk. Under den stationära fasen dör bakterier med ungefär samma hastiget, som de återbildas vid celldelningarna och i deklinationsfasen dör bakterierna snabbare än de återbildas.
När man bestämmer en tillväxtkurva vill man oftast kunna följa tillväxten av bakterier i realtid för att veta när man ska göra t.ex. en tillsats av någon substans eller när odlingen bör avbrytas. Den snabbaste och enklaste metoden är att regelbundet ta ut prover från kulturen och bestämma ljusspridningen genom spektrofotometri. Med en spektrofotometer mäter man normalt absorbans vid olika våglängder, men i en suspension (av t.ex. bakterier) kan man även mäta ljusspridning (OD = optical density). Ljusspridningen är proportionell mot antal bakterier per ml och med hjälp av en standardkurva kan man bestämma antalet bakterier i absoluta tal.
Uppdaterad: 2025-05-23.
Bakterietoxiner är viktiga virulensfaktorer och de brukar indelas i exotoxiner (exo = ute) och endotoxiner (endo = inne). Exotoxiner syntetiseras och utsöndras aktivt, medan endotoxiner utgör en del av bakterien och frigörs först när bakterierna dör och lyserar. De flesta endotoxiner är lokaliserade till bakteriens hölje och vanligen avses endast lipopolysackarider (LPS) i yttermembranet hos gramnegativa bakterier (se även lipopolysackarid ovan). Endotoxiner är värmestabila.
Exotoxiner utgörs vanligen av värmekänsliga (värmelabila) proteiner, men kan också vara värmestabila polypeptider. Exotoxiner kan bildas av både grampositiva och gramnegativa bakterier. Bland de bakteriella exotoxinerna hittar man några av naturens mest potenta toxiner. Dödlig dos för människa av t.ex. botulinumtoxinet är endast 1-2 ng (1 ng = 0,000 000 001 g) per kg kroppsvikt vid intrevenöst upptag. Detta innebär att botulinumtoxinet är ca. 6 000 000 gånger giftigare än skallerormsgift.
Exotoxiner kan i sin tur indelas efter t.ex.:
De olika klassificeringsmodellerna är olyckligtvis en källa till missförstånd eftersom ett och samma toxin kan få olika namn beroende på vilken klassificeringsmodell som har tillämpats. Botulinumtoxinet t.ex. är både ett neurotoxin och ett AB-toxin. Man använder ofta förkortningar och botulinumtoxinet t.ex. kan betecknas BoNT.
En toxoid är ett proteintoxin, som har denaturerats (genom t.ex. värmebehandling eller kemisk behandling) och därför förlorat sin toxicitet. En toxoid har däremot kvar sina antigena egenskaper och kan därför användas i vacciner.
LPS finns ej hos grampositiva bakterier, men förutom LPS finns det några andra endotoxiner och ett exempel är cryproteinet hos Bacillus thuringiensis, som är ett s.k. δ-endotoxin. Endotoxiner är i allmänhet mycket mindre toxiska än exotoxiner.
Bakterier har komplicerade sekretionssystem for att kunna exportera proteiner till omgivningen eller till cytoplasman hos värdorganismens celler. Proteiner som ska exporteras är ofta toxiner. Information om olika sekretionssystem finns också i termlitan. Se Sekretionssystem, bakteriella.
Uppdaterad: 2019-01-25.
Exempel på bakteriesläkten och arter, som kan övergå till VBNC-tillståndet: Aeromonas, Burkholderia, Campylobacter, Enterobacter, Entercoccus, Escherichia coli, Francisella, Helicobacter, Klebsiella, Legionella, Listeria, Mycobacterium, Pasteurella, Pseudomonas, Salmonella, Serratia, Shigella, Streptococcus och Vibrio.
Uppdaterad: 2013-02-26.
Virulens är inte synonymt med patogenicitet (se även patogenicitet ovan), utan beskriver graden av skada, som den patogena organismen har orsakat värddjuret. En högvirulent bakterie är mycket smittsam och/eller ger kraftiga symptom. Infektionsdosen (ID) är alltså lägre för en högvirulent än för en lågvirulent patogen.
Virulensfaktorer är de komponenter, som bakterien tillverkar för att kunna:
Virulensgener är de gener, som kodar för virulensfaktorer eller komponenter, som behövs för syntes av virulensfaktorer.
Uppdaterad: 2025-01-08.
.")
Avibacterium paragallinarum, odlad på nötblodagar tillsammans med ett stryk av Staphylococcus aureus. A. paragallinarum "ammar" dvs. växer bättre och ger större kolonier i närheten av stryket av Staphylococcus aureus, som lyserar blodkropparna, vilket gör att V-faktorn frisätts.
Bild: Ingrid Hansson & Karl-Erik Johansson (BVF, SLU). - Klicka på bilden för att förstora den.
Inledning
Vissa bakterier producerar substanser, som andra bakterier behöver för tillväxt. Exempel på sådana substanser är X-faktorn och V-faktorn. X-faktorn, som är värmestabil, kan utgöras av protoporfyrin IX, hemin eller andra järn-innehållande porfyriner och finns både i röda blodkroppar samt extracellulärt i blod. X-faktorn kallas även för Stuart-Prowers faktor och utgör både en koagulationsfaktor (i koagulationssammanhang) och en tillväxtfaktor (i bakteriologiska sammanhang). V-faktorn, som är värmekänslig, kan utgöras av nikotinamid-adenin-dinukleotid (NAD) eller nikotinamid-adenin-dinukleotidfosfat (NADP) och finns framför allt intracellulärt i röda blodkroppar. X- och V-faktorberoende kan påvisas antingen genom ett X- och V-faktortests (även kallat amningstest) eller genom ett disktest och resultatet kan ge värdefull information vid identifiering av bakterier.
Med amningstestet undersöker man hur en viss bakterier växer på blodagar och på ett minimalmedium (t.ex. TSA) i närheten av ett stryk av en bakterie, som producerar V-faktor under tillväxten. Först inoculeras de båda plattorna med den bakterie, som ska undersökas, och därefter gör man ett stryk av Staphylococcus aureus på båda plattorna. S. aureus producerar alltså V-faktor. Därefter inkuberas plattorna i 5-10% CO2 vid 37 °C under 2-3 dygn. Bakterier, som bara kräver V-faktor, kommer att bilda större kolonier (s.k. satellitkolonier) runt S. aureus-stryket på båda plattorna medan bakterier, som kräver både V- och X-faktor, kommer bara att växa på blodagar-plattan och ge större satellitkolonier runt S. aureus-stryket.
Med ett disktest kan man också undersöka om en bakterie kräver V-faktorn eller både X- och V-faktorn. Testet utförs då med tre filterpapperdiskar indränkta med respektive V faktor, X-faktor samt en blandning av X- och V-faktor. Diskarna ska sedan appliceras på en agarplatta där bakterien, som ska undersökas, har inokulerats. Denna agarplatta (t.ex, TSA) får inte innehåller X- eller V-faktor. Bakterien kommer då att kunna växa ut som satellitkolonier nära respektive disk med den eller de tillväxtfaktorer, som bakterien i fråga behöver.
Hematinagar används för att odla kräsna bakterier och det innehåller blod, som har hemolyserats genom uppvärmning. X-faktorn, som är värmestabil, har alltså frigjorts från de röda blodkropparna. V-faktorn, som är värmelabil, har delvis förstörts, trots att NAD+-nukleosidaset (NADas), som finns i blod, har inaktiverats av värmen.
X-faktorn (hemin) behövs för syntes av cytokrom C och andra järninnehållande respiratoriska enzymer. V-faktorn (NAD och NADP) behövs för reduktions-oxidations (redox) processer i cellmetabolismen.
Uppdaterad: 2024-11-04.